








En la década de los 90 se descubrió un nuevo sistema de comunicación célula-célula, que consiste en la liberación de vesículas cargadas con partículas bioactivas (proteínas, mRNA, miRNA, metabolitos, etc.) en el espacio extracelular. Este tipo de comunicación se ha conservado durante la evolución, hecho que justificaría que la mayoría de los tipos celulares puedan generarlas. Estas vesículas extracelulares (VE) pueden regular diversos procesos fisiológicos, así como el desarrollo y progresión de enfermedades. En los últimos años se ha extendido el estudio de las VE generadas principalmente por células madre adultas o embrionarias, células sanguíneas, células del sistema inmune y nervioso, así como células tumorales. El análisis de VE en fluidos corporales ha sido utilizado como herramienta de diagnóstico en cáncer y recientemente para distintas enfermedades renales. Sin embargo, en esta revisión pretendemos analizar la importancia, función y posible aplicación clínica de las VE generadas por células madre en enfermedades renales y en trasplantes.
A new cell-to-cell communication system was discovered in the 1990s, which involves the release of vesicles into the extracellular space. These vesicles shuttle bioactive particles, including proteins, mRNA, miRNA, metabolites, etc. This particular communication has been conserved throughout evolution, which explains why most cell types are capable of producing vesicles. Extracellular vesicles (EVs) are involved in the regulation of different physiological processes, as well as in the development and progression of several diseases. EVs have been widely studied over recent years, especially those produced by embryonic and adult stem cells, blood cells, immune system and nervous system cells, as well as tumour cells. EV analysis from bodily fluids has been used as a diagnostic tool for cancer and recently for different renal diseases. However, this review analyses the importance of EVs generated by stem cells, their function and possible clinical application in renal diseases and kidney transplantation.
La aplicación de terapias celulares para frenar la progresión de enfermedades renales es una aproximación muy prometedora debido a su capacidad inmunomoduladora y regenerativa1–5. El efecto protector de células madre mesenquimales (MSC) no es debido a su transdiferenciación, sino al impacto de su actividad en el tejido dañado1. Sin embargo, antes de llegar a la práctica clínica de rutina existen una serie de aspectos de seguridad que deben ser analizados con más detenimiento: la posibilidad de rechazo inmunológico del receptor, la estabilidad genética de las células, la mala diferenciación a largo término y la probabilidad de transferencia viral6–8. Por ello, se ha impulsado el estudio de los mecanismos de acción asociados a su capacidad protectora y regenerativa para diseñar una terapia alternativa libre de células. Así, se describió que el secretoma o medio condicionado de las MSC puede ejercer el mismo efecto protector que las MSC sobre los daños tisulares y contribuir a la inmunomodulación del estado inflamatorio9–13. El análisis del medio condicionado puso de relevancia la presencia de factores de crecimiento, citocinas y de vesículas extracelulares (VE). Estas últimas pueden transferir proteínas, lípidos y material genético a las células residentes en el tejido dañado. Las VE contribuyen activamente a la capacidad terapéutica de las MSC y, en particular, a la reprogramación de las células residentes mediante la transferencia horizontal de mRNA y miRNA9,14–25. Tras demostrarse que las VE poseen la misma capacidad terapéutica que las MSC, se está promoviendo a las VE como terapia libre de células por su mayor seguridad sanitaria26.
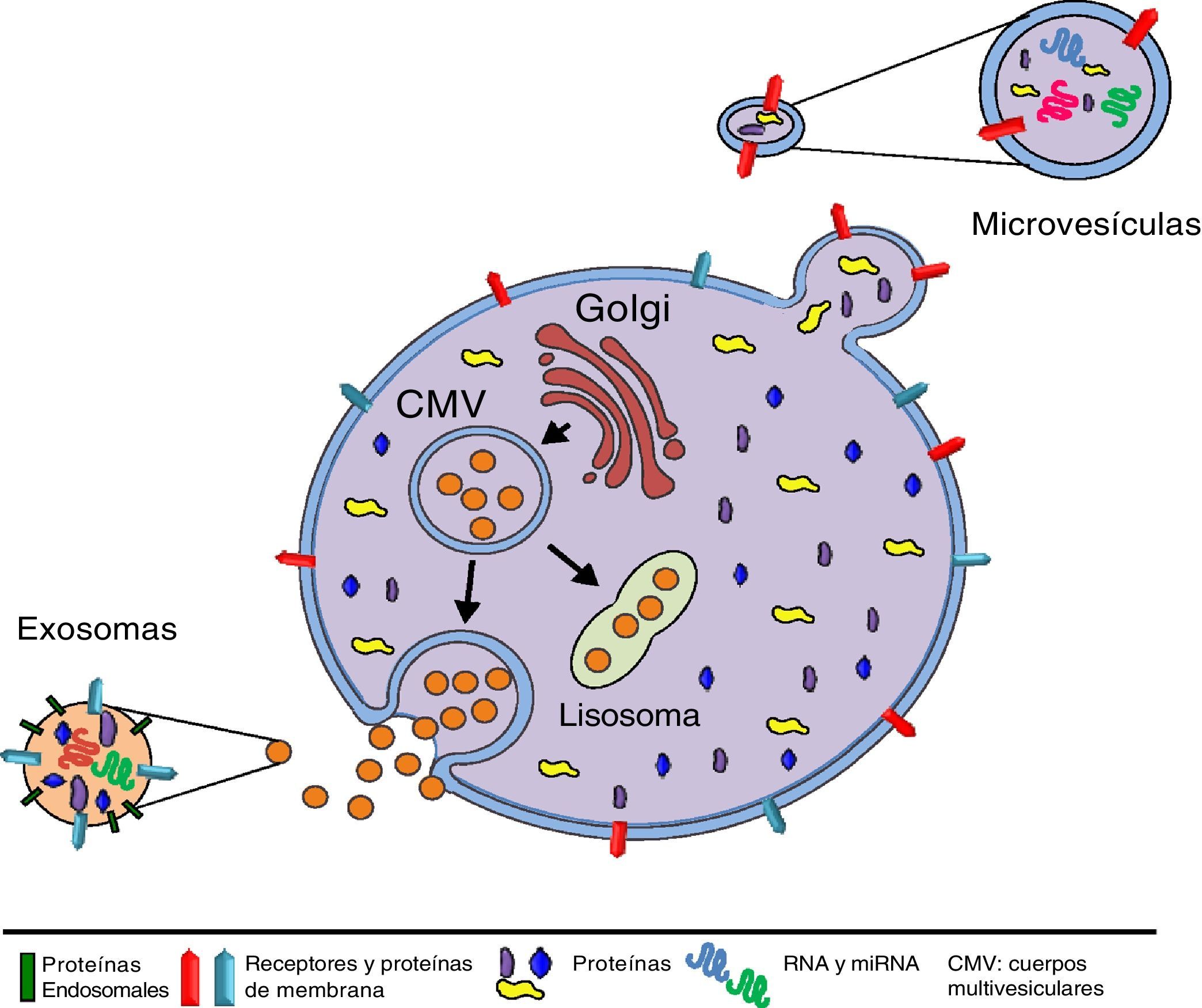
La comunicación célula-célula mediante VE es un mecanismo que se ha conservado a lo largo de la evolución tanto en células eucariotas como en procariotas27. Desde su descubrimiento hace 30 años28, se ha demostrado que las VE son producidas por una gran variedad de tipos celulares: células sanguíneas, dendríticas, endoteliales y epiteliales, células del sistema nervioso, células madre adultas y embrionarias e incluso células tumorales.
Las VE están formadas por una membrana lipídica y pueden transmitir señales biológicas reguladoras mediante la transferencia de proteínas de membrana y citosólicas, lípidos, mRNA, miRNA, DNA mitocondrial y DNA genómico que regulan diversos procesos fisiológicos, así como en el desarrollo y progresión de enfermedades29–34. En el contexto fisiológico, todas las células pueden producir VE como mecanismo habitual de comunicación paracrina/endocrina, si bien en el momento en que tiene lugar un daño la producción de VE se ve aumentada y el contenido vesicular modificado para alertar a las células adyacentes, progenitoras y del sistema inmunitario. Todo ello, para intentar restablecer la homeostasis del tejido dañado. Solamente las células progenitoras y las MSC pueden generar VE con capacidad protectora o regeneradora intrínseca.
En cuanto a la progresión de enfermedades, se ha demostrado que el microambiente define el contenido de las VE. Concretamente en la arterioesclerosis, las células del endotelio vascular sometidas a un ambiente de estrés mediado por calcio generan VE que promueven la mineralización del tejido35.
En el contexto tumoral, se ha postulado que las células progenitoras al sufrir mutaciones pueden ser el origen de las cancer stem cell36, las cuales producen VE implicadas en el desarrollo y progresión del tumor. Estas VE promueven angiogénesis37, permiten escapar de la inmunovigilancia38, inducen la eliminación de moléculas terapéuticas que activan la apoptosis39 y participan activamente en la degradación de la matriz extracelular necesaria en la metástasis40. Actúan como efectores paracrino/endocrino transportando moléculas bioactivas entre las células del microambiente, o bien, de forma remota mediante su transporte en fluidos biológicos41.
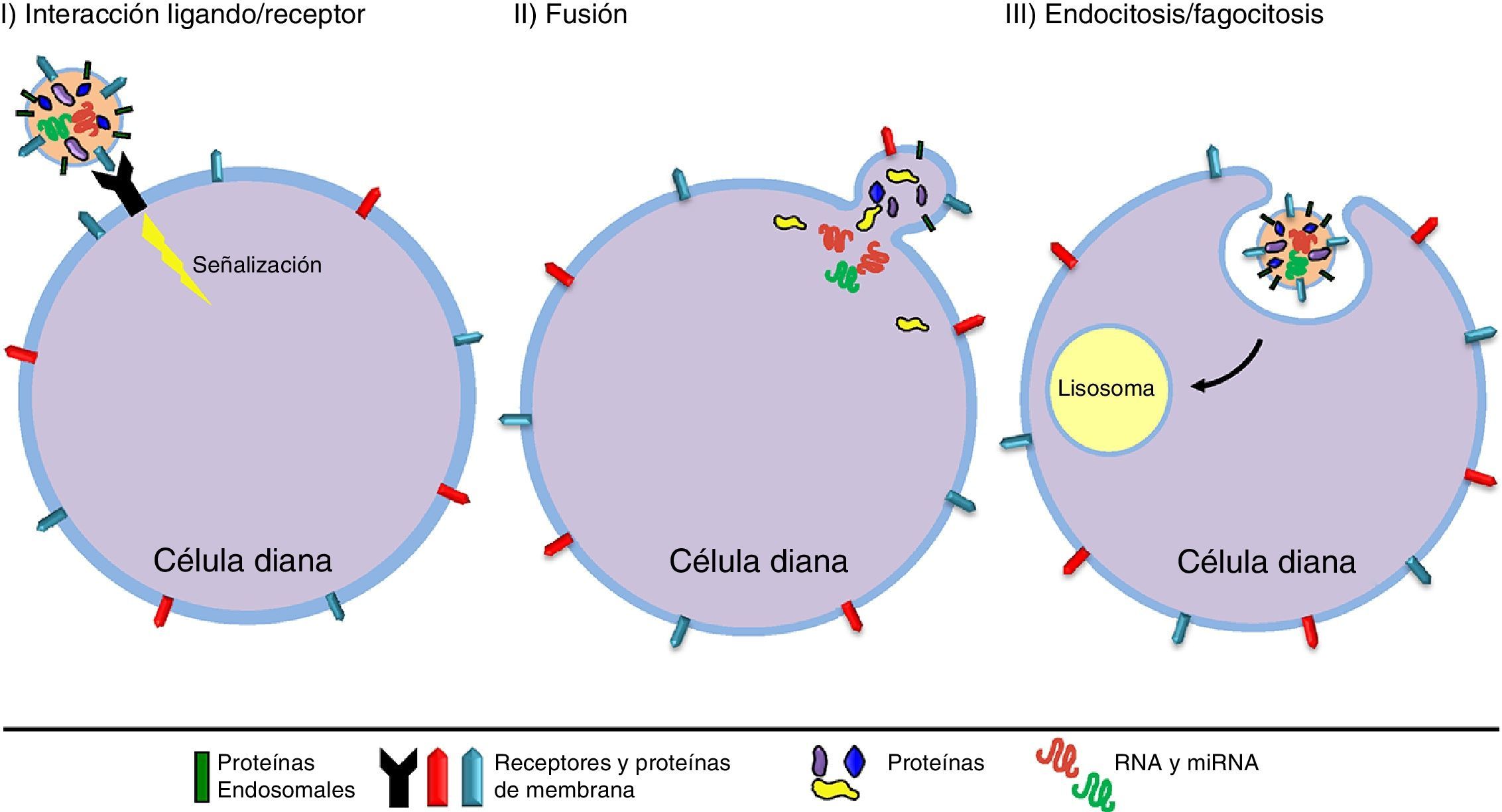
El origen y tamaño de las VE nos permite diferenciar entre exosomas (EX) y microvesículas (MV) (fig. 1). Los EX son vesículas de tamaño pequeño (70-150nm) y tienen un origen endosomal, por consiguiente, su membrana está enriquecida con colesterol, ceramidas y esfingolípidos, y su contenido corresponde al existente en el compartimento endosomal. En cambio, las MV tienen un tamaño mayor (150-1.000nm), se generan como resultado de evaginaciones de la membrana plasmática y su composición depende del tipo celular del que proceden42,43 (tabla 1). Las VE pueden actuar mediante 3 mecanismos (fig. 2): I) Activando una vía de señalización de la célula diana por su adhesión con elevada especificidad en la superficie de la célula diana (sin fusión de membrana), mediante las moléculas de adhesión y los receptores presentes en la superficie celular44. II) Transfiriendo mRNA, miRNA, proteínas y moléculas de señalización, mediante la fusión de la membrana45. III) Incorporando su contenido mediante endocitosis en las células diana y procesado de su contenido en el compartimento endosomal46.
Características de las vesículas extracelulares
| Exosomas | Microvesículas | |
|---|---|---|
| Tamaño en nm | 70-150 | 150-1.000 |
| Composición lipídica | Poca exposición de fosfatidilserina. Ácido lisobifosfatídico. Colesterol y ceramida | Elevada exposición de fosfatidilserina. Colesterol |
| Marcadores proteicos | Alix, TSG101, HSC70, CD63, CD81, CD9 | Selectinas, integrinas, CD40, metaloproteinasas |
| Origen | Cuerpo multivesicular | Membrana plasmática |
| Mecanismos de secreción | Exocitosis del cuerpo multivesicular | Evaginación de la membrana plasmática |
| Composición | Proteínas, mRNA y miRNA | Proteínas, mRNA y miRNA |
En la actualidad, las VE se están usando como biomarcadores en orina de diferentes enfermedades renales e incluso para el rechazo del injerto renal47–50. Sin embargo, el objetivo de esta revisión es analizar la importancia, función y posible aplicación de las VE en enfermedades renales como terapia alternativa o bien terapia complementaria al uso de los inmunosupresores en el caso del trasplante renal. Por ello, analizaremos diferentes modelos in vitro e in vivo de daño renal en los que las VE procedentes de MSC han inducido regeneración15,51–57.
Insuficiencia renal agudaLa insuficiencia renal aguda (IRA) se caracteriza por una pérdida en horas o días de la función excretora renal, con la acumulación de productos del metabolismo nitrogenado como la creatinina y la urea. Aunque las causas son controvertidas, las más comunes son una lesión por isquemia y reperfusión (I/R) o la exposición a agentes nefrotóxicos que conducen a una necrosis tubular aguda. Estos insultos a nivel tubular conllevan la generación de mediadores inflamatorios que promueven la vasoconstricción y favorecen el proceso inflamatorio acompañado de la infiltración de neutrófilos secretores de especies reactivas de oxígeno, proteasas y mieloperoxidasas que agravan el daño renal. La IRA se considera una de las mayores causas de morbimortalidad en los pacientes hospitalizados, por ello hay tantos estudios que buscan biomarcadores que permitan un diagnóstico precoz, o bien que establezcan nuevas terapias1,58–60.
El efecto protector y regenerativo de las MSC en modelos animales de la IRA está ampliamente documentado, sin embargo, en los últimos años han surgido importantes evidencias que demuestran que su efecto protector se debe a la acción paracrina y no a la posible transdiferenciación1,9,17–25. Este efecto paracrino incluye una acción proliferativa, antiapoptótica e inmunomoduladora. Concretamente, en el contexto del órgano dañado se genera un microambiente rico en citocinas como el interferón gamma (IFNγ) y el factor de necrosis tumoral alfa (TNFα), que estimulan las MSC e inducen la secreción de diferentes factores tróficos, de crecimiento, citocinas y VE61–63.
El efecto protector de las VE se ha analizado en varios modelos de IRA en animales de experimentación: daño renal inducido por glicerol15,16,64,65, cisplatino54, gentamicina55, I/R52,66–70, obstrucción unilateral del uréter71 y nefrectomía 5/6 a 7 días poscirugía72 (tabla 2). Estos modelos de IRA se caracterizan por presentar daño tubular con un elevado componente inflamatorio asociado a un incremento del infiltrado intersticial, apoptosis y necrosis tubular. Este microambiente induce la selección de las VE del mismo modo que sucede con las MSC. La acumulación de VE a nivel tisular podría verse favorecida por el aumento de la permeabilidad que presentan los tejidos dañados64. La internalización de la VE depende de la presencia de receptores celulares (CRCX4) y de las moléculas de adhesión (CD44 y CD29), estas últimas localizadas tanto en las membranas de las VE como en las MSC21. Las VE más extensamente estudiadas han sido las generadas por las células humanas mesenquimales de médula ósea (BM-MSC), pero también se han llevado a cabo estudios con VE de otros orígenes como las células madre humanas de hígado, células madre mesenquimales de sangre de cordón umbilical, células humanas mesenquimales procedentes de la gelatina de Wharton, células progenitoras endoteliales circulantes (EPC) y células madre mesenquimales derivadas del riñón. Todas estas VE aceleran la recuperación del daño renal agudo de la misma manera que lo hacen las células productoras. Esta renoprotección se caracteriza por una mejora de los parámetros bioquímicos renales, con la disminución de las concentraciones de los niveles de nitrógeno ureico en sangre y de la creatinina en suero, y una mejora de las lesiones histológicas.
Resumen de los modelos animales de insuficiencia renal aguda donde se aplica una terapia con VE
| Modelo | Origen celular de las VE | Tipo de VE | Vía de administración | Capacidad terapéutica | Referencia |
|---|---|---|---|---|---|
| Glicerol | BM-MSC | MV | Intravenosa | Recuperación morfológica y funcional mediante transferencia de mRNA y miRNA | 16 |
| HLSC | MV | Intravenosa | Disminución de la necrosis y la proliferación tubular | 65 | |
| BM-MSC | MV | Intravenosa | Recuperación morfológica y funcional | 64 | |
| BM-MSC | MV | Intravenosa | Inducción de la proliferación | 15 | |
| Isquemia-reperfusión | EPC | EX | Intravenosa | Disminución del efecto apoptótico y proinflamatorio Presencia de transcritos proangiogénicos | 70 |
| BM-MSC | MV | Intravenosa | Inducción de la proliferación e inhibición de la apoptosis. Efecto proangiogénico | 68 | |
| WJ-MSC | MV | Intravenosa | Inducción de la proliferación e inhibición de la apoptosis Efecto antiinflamatorio | 69 | |
| UCB-MSC | MV | Intravenosa | Modulación de la células T. | 67 | |
| EPC | MV | Intravenosa | Inducción de la proliferación, inhibición de la apoptosis y del infiltrado leucocitario. Efecto proangiogénico | 66 | |
| BM-MSC | MV | Intravenosa | Inducción de la proliferación, inhibición de la apoptosis y del infiltrado leucocitario | 52 | |
| Cisplatino | UCB-MSC | EX | Inyección en la capsula renal | Disminución de la apoptosis, necrosis de las células tubulares y estrés oxidativo | 73 |
| BM-MSC | MV | Intravenosa | Mejoraría de la función renal y de la supervivencia | 54 | |
| Gentamicina | BM-MSC | EX | Intravenosa | Inducción de la proliferación e inhibición de la apoptosis y de la necrosis. Efecto antiinflamatorio | 55 |
| Obstrucción unilateral del uréter | BM-MSC | MV | Intravenosa | Disminución del infiltrado linfocitario, inflamación tubular y necrosis | 71 |
| Nefrectomía 5/6 | BM-MSC | MV | Intravenosa | Disminución de la fibrosis, infiltrado linfocitario intersticial y disminución o ausencia de atrofia tubular | 72 |
Los mecanismos de acción de las VE se han analizado en diferentes modelos in vitro que incluyen el uso de células epiteliales tubulares renales, células endoteliales y células mononucleares de sangre periférica. En todos ellos se ha observado que las VE dan lugar a una disminución de la expresión de moléculas inflamatorias, estimulan la proliferación e inhiben la apoptosis. El análisis proteómico de la MV derivadas de hUCB-MSC demostró la presencia de proteínas que ejercen un efecto protector del epitelio endotelial y tubular. Concretamente, hallaron galectina-1 y 3, mediadores en la regulación que ejercen las MSC sobre las células T, 2 marcadores de MSC, CD73 y CD90, asociados con su capacidad inmunosupresora y algunos elementos de la vía de complemento como el CD59, C5, C3 y C4A. Además, apolipoproteínas como ApoA2, ApoA4 y ApoC3, que se ha visto que tienen un efecto protector en el endotelio vascular en diversas condiciones patológicas, y proteínas transportadoras de lípidos como SCP2 y FABP667. Este contenido proteico se ve modificado radicalmente si las células reciben un estímulo proinflamatorio (IFNγ), que reduce considerablemente la capacidad protectora de las MV67.
El daño oxidativo, característico de la enfermedad renal inducida por la administración de cisplatino, se ve reducido por la acción de los EX de hUCB-MSC, que disminuyen la formación de productos oxidativos, 8-HdG y MDA, e incrementan los niveles de GSH. A su vez, los EX promueven la proliferación celular al activar ERK1/273.
El daño agudo induce la apoptosis en la célula tubular renal. Para estudiar el impacto de las VE sobre la apoptosis se han utilizado los cultivos de células epiteliales tubulares renales en presencia de cisplatino. Las MV de BM-MSC detienen la activación de genes relacionados con la parada del ciclo celular como GADD45A, de la apoptosis como Bcl-10, CASP-1, CASP-8, LTA, TP73 y CASP-10 y disminuyen la expresión de genes antiapoptóticos como Bcl2, Bcl-XL, Akt1 y TRAF2. Además, se observa un aumento de la síntesis de factores renoprotectores como el factor de crecimiento del hepatocito y la proteína estimulante de los macrófagos. El mRNA de estos factores no forma parte del contenido de las MV, lo que indica que la activación de las vías metabólicas que desencadena un fenotipo regenerativo debe inducirse por factores proteicos15,54.
Además del contenido proteico, el material genético (mRNA y miRNA) incluido en las VE también puede modificar los patrones de expresión normales de las células residentes mediante su transferencia horizontal74. En el caso de las MV derivadas de las células madre mesenquimales del riñón, presencia de mRNA codificante para el factor de crecimiento del endotelio vascular (VEGF-A), el factor de crecimiento insulínico tipo 1 y el factor básico de crecimiento del fibroblasto estimulan las células endoteliales promoviendo la angiogénesis68, mientras que el estudio de las MV derivadas de EPC denota que la presencia de miR-126 y miR-296 son clave para estimular la angiogénesis. El análisis del transcriptoma de las MSC y sus MV puso de relevancia la presencia de transcritos relacionados con la diferenciación celular, el control de la transcripción, proliferación y de la regulación del sistema inmune. También se observaron transcritos relacionados con la oxidación de los ácidos grasos, glucólisis, gluconeogénesis y de la generación de cuerpos cetónicos, procesos fundamentales para la citoprotección de las células tubulares renales en procesos como la IRA16. La ratificación de la importancia de los mRNA y miRNA sobre el efecto protector de las VE se ha demostrado mediante distintas estrategias: usando ribonucleasasas para eliminar todo el mRNA y miRNA, deplecionando específicamente los miR-126 y el miR-296 con miRNA antagónicos, o bien deplecionado todos los miRNA mediante la eliminación de los genes Dicer o Drosha, esenciales para la producción y maduración de los miRNA, en las células generadoras de EV14–16.
Actualmente hay varios estudios clínicos en marcha o a punto de iniciarse que pretenden indagar sobre la eficacia y seguridad de la aplicación de las MSC en pacientes con insuficiencia renal aguda (tabla 3). Sin embargo, el uso de VE para solventar enfermedades renales agudas humanas todavía no se ha iniciado.
Ensayos clínicos realizados con células madre para tratar la insuficiencia renal aguda
| Patología | Número Trial | Título | Tipo celular | Estado actual |
|---|---|---|---|---|
| Insuficiencia renal aguda | NCT01275612 | MSC in cisplatin-induced acute renal failure in patients with solid organ cancers (CIS/MSC08) | MSC | Seleccionando |
| Insuficiencia renal aguda | NCT00733876 | Allogeneic multipotent stromal cell treatment for acute kidney injury following cardiac surgery | BM-MSC | Completo |
| Insuficiencia renal aguda | NCT01602328 | A study to evaluate the safety and efficacy of AC607 for the treatment of kidney injury in cardiac surgery subjects (ACT-AKI) | AC607 Allogeneic BM-MSC | Terminado |
| Hipertensión renovascular | NCT02266394 | Hypoxia and inflammatory injury in human renovascular hypertension | MSC | Seleccionando |
El incremento de la prevalencia de la enfermedad renal crónica en la población adulta hace muy necesario identificar terapias que frenen o reviertan su progresión hasta el fallo renal terminal.
Las MSC se han utilizado en distintos modelos animales en los que se simula la enfermedad renal crónica: modelos de masa renal reducida (nefrectomía 5/6)2,3,75–78, poliquistosis renal5, nefropatía diabética79,80, inducción de glomerulosclerosis por adriamicina4, estenosis aterosclerótica81 o el modelo de enfermedad renal crónica inducida por cisplatino82. En el caso del modelo de nefrectomía 5/6, en todos los estudios las BM-MSC mejoran la función renal y reducen la fibrosis. Esta mejoría está asociada a una reducción de la progresión de la glomeruloesclerosis2,3,75 y de la expresión de interleucina-6 (IL-6) y TNFα, mientras que la expresión de IL-4 e IL-10 se ve aumentada76. También se observa una reducción de la expresión del factor de crecimiento del endotelio vascular, p21 y del antígeno nuclear de células en proliferación77, y la formación de nuevo epitelio mediante la activación de Pax-1, factor básico de crecimiento del fibroblasto, la proteína morfogénica del hueso (BMP-7) y Tie-278. En el modelo de poliquistosis renal en rata, se ha demostrado que las BM-MSC permiten la mejora de la densidad vascular y, por consiguiente, la función renal5. En distintos estudios en los que se analiza la nefropatía diabética, la administración de BM-MSC de origen murino o humano ha demostrado la reversión de la hiperglucemia y glucosuria inducida por la administración de estreptozotocina79,80. En el modelo de inducción de glomerulosclerosis por adriamicina, las MSC pueden desplazarse hasta el riñón dañado y proveer de factores de supervivencia que preservan la viabilidad de los podocitos mientras reducen la inflamación y esclerosis glomerular4. En el modelo de estenosis aterosclerótica en la arteria renal inducida en cerdo, se observa un daño renal crónico caracterizado por una extensa inflamación, apoptosis, estrés oxidativo, pérdida de microvasculatura, fibrosis y glomerulosclerosis. La administración de MSC de tejido adiposo tras una angioplastia transluminal percutánea renal permite mejoran la función renal a través de la revascularización del riñón81. En el modelo de enfermedad renal crónica inducida por cisplatino en primates no humanos se ha demostrado que el uso preventivo de las BM-MSC autólogas permite retrasar la progresión de la fibrosis intersticial, pero no es capaz de revertir el daño establecido82.
En clínica se ha visto que la administración de 3 dosis de BM-MSC alogénicas a un paciente con glomerulosclerosis focal y segmentaria recurrente tras el trasplante renal permitió frenar la proteinuria y mantener una función renal estable, y evitar el tratamiento convencional que implicaba plasmaférisis semanal83.
La existencia de todos estos hallazgos y muchos otros han dado información suficiente para que se empiecen a realizar ensayos clínicos para evaluar la eficacia y seguridad de las MSC (tabla 4).
Ensayos clínicos realizados con células madre para tratar la enfermedad renal crónica
| Enfermedad | Número Trial | Título | Tipo celular | Estado actual |
|---|---|---|---|---|
| Enfermedad renal crónica | NCT02195323 | (BM-MSC) in patients with chronic kidney disease (CKD) | Autologous BM-MSC | Completo |
| Enfermedad renal crónica | NCT01876017 | Safety and efficacy of BMMNC in patients with chronic renal failure | BM-mononuclear stem cell (BM-MNC) | Desconocido |
| Enfermedad renal crónica | NCT01453816 | Study to assess the safety and effects of autologous adipose-derived stromal cells delivered in patients with renal failure | Autologous adipose-MSC | Desconocido |
| Poliquistosis renal-enfermedad renal crónica | NCT02166489 | Mesenchymal stem cells transplantation in patients with chronic renal failure due to polycystic kidney disease | MSC | Completo |
| Nefropatía diabética-enfermedad renal crónica | NCT02585622 | Novel stromal cell therapy for diabetic kidney disease | Novel stromal cell | Todavía no ha empezado la selección |
En los últimos años algunos grupos han demostrado extensamente el efecto positivo de las VE en modelos de insuficiencia renal aguda, sin embargo, su aplicación en modelos crónicos ha sido nula. Los únicos estudios para analizar el impacto de las VE en el daño crónico han consistido en la prolongación del seguimiento de un modelo de daño agudo en animales tratados mediante I/R donde se ha observado que las VE producidas por EPC, así como aquellas producidas por BM-MSC, simulan el efecto de las MSC y previenen el daño agudo y también el crónico inducido por la I/R14,52. Según estos estudios, los micro-RNA —miR-126 y miR-296— son responsables del buen resultado de las VE de EPC14, mientras que el contenido de RNA de las VE producidas por BM-MSC es el responsable de la activación de las células diana52.
Fallo renal terminal: terapia celular y sus derivadosLa progresiva pérdida de función renal en la enfermedad renal crónica terminal se asocia a la activación del sistema inmune, marcada por una inflamación renal y sistémica. En dicha inflamación se observa activación del sistema del complemento, monocitos, macrófagos y quimiocinas84. El trasplante es la mejor terapia sustitutiva para prolongar la supervivencia de los pacientes con enfermedad renal crónica terminal, pero el daño isquémico es una de las causas del retraso de la función del injerto, el cual se ha asociado a un incremento de episodios de rechazo agudo y a la reducción de la supervivencia del injerto a largo plazo85. Los factores de riesgo más importantes para la progresiva pérdida de función renal en riñones trasplantados son el daño por I/R y la proteinuria86. A pesar de las nuevas estrategias inmunosupresoras, los resultados a largo plazo no han mejorado en la última década, debido al desarrollo de disfunción crónica del injerto y a la mortalidad de los pacientes con injertos funcionales. Esta última es, principalmente, por causas cardiovasculares y neoplasias malignas87–91.
El uso prolongado de los inmunosupresores conlleva efectos secundarios leves que en algunos casos puede contribuir a la aparición de enfermedades más graves, entre ellas, diabetes y neoplasias. Por ello, es necesaria la búsqueda de terapias alternativas que permitan reducir el uso de inmunosupresores e, idealmente, induzcan la tolerancia del injerto. En ese sentido, el uso de MSC o bien de células reguladoras ha sido un punto de partida muy interesante. En distintos estudios con animales de experimentación se ha demostrado el efecto beneficioso de las terapias celulares en el trasplante renal, que evita el daño inducido por I/R, la fibrosis intersticial, atrofia tubular y el rechazo agudo92,93.
Actualmente hay varios ensayos clínicos para testar la eficacia y seguridad de las terapias celulares en el trasplante renal, incluyendo células reguladoras y células madre de distintos orígenes (tabla 5).
Ensayos clínicos realizados con terapia celular para tratar el trasplante renal
| Patología | Número Trial | Título | Tipo celular | Estado actual |
|---|---|---|---|---|
| Trasplante renal | NCT02085629 | Mreg (The ONE Study) | Donor M reg (Mreg_UKR) | Seleccionando |
| Trasplante renal | NCT02129881 | TregUK (The ONE Study) | Autologous regulatory T Cell Product | Seleccionando |
| Trasplante renal | NCT02371434 | nTreg Trial (The ONE Study) | Autologous CD4+CD25+FoxP3+ natural regulatory.T cells | Inclusión por invitación |
| Trasplante renal | NCT02091232 | T-regulatory cells in kidney transplant recipients (The ONE Study) | T regulatory cell | Seleccionando |
| Trasplante renal | NCT02244801 | darTreg (The ONE Study) | Donor-alloantigen-reactive regulatory T Cell | Seleccionando |
| Trasplante renal | NCT02252055 | ATDC Trial (ONEatDC) | Autologous tolerogenic dendritic cells (ATDCs) | Seleccionando |
| Trasplante renal | NCT01446484 | Treatment of children with kidney transplants by injection of CD4+CD25+FoxP3+ T cells to prevent organ rejection | CD4+CD25+CD127low FoxP3+ T regulatory cells injection | Desconocido |
| Trasplante renal | NCT02560220 | MIC cell therapy for individualized immunosuppression in living donor kidney transplant recipients | Mitomycin C-induced PBMC | Seleccionando |
| Trasplante renal | NCT02057965 | Mesenchymal stromal cell therapy in renal recipients | Autologous BM-MSC | Seleccionando |
| Trasplante renal | NCT02176434 | Pilot feasibility study of combined kidney and hematopoietic stem cell transplantation to cure end-stage renal disease | Hematopoietic stem cell | Seleccionando |
| Trasplante renal o hepático | NCT01429038 | Infusion of third-party mesenchymal stem cells after renal or liver transplantation. A phase I-II, open-label, clinical study | Third party MSC | Seleccionando |
| Trasplante renal | NCT00658073 | Induction therapy with autologous mesenchymal stem cells for kidney allografts | Autologous MSC | Completo |
| Trasplante renal | NCT02387151 | Neptune | Allogeneic MSC | Seleccionando |
| Trasplante renal | NCT02565459 | MSC in renal recipients to induce tolerance in recipients of kidney transplants from deceased donors | Third-party BM-MSC | Seleccionando |
| Tolerancia | NCT00183248 | Pilot study using donor stem cells and campath-1H to induce renal transplant tolerance (ITN022ST) | Donor BM-stem cells | Completo |
| Tolerancia | NCT00752479 | MSC and kidney transplant tolerance | MSC under basiliximab/low dose RATG | Terminado |
| Tolerancia | NCT02012153 | aMSC to induce tolerance in living-donor kidney transplant recipients | Autologous MSC | Seleccionando |
| Rechazo crónico | NCT02563340 | Effect of BM-MSC on chronic AMR after kidney transplantation | BM-MSC | Todavía no ha empezado la selección |
| Función retardada | NCT02563366 | Effect of BM-MSC on early graft function recovery after DCD kidney transplant | BM-MSC | Todavía no ha empezado la selección |
| Función retardada | NCT02561767 | Effect of BM-MSC in DCD kidney transplantation | BM-MSC | Todavía no ha empezado la selección |
El uso de las VE en los modelos experimentales de trasplante renal es muy anecdótico hasta el momento. Solo hay un estudio en el que se analiza el papel de las VE isogénicas producidas por BM-MSC, las cuales consiguen reducir la respuesta innata mediante la reducción de células natural killer infiltradas en el injerto, así como una reducción de TNFα; sin embargo, no consiguen suprimir la respuesta inmune adaptativa94. En ese sentido, la ayuda a la investigación que nos concedió la Sociedad Española de Nefrología en 2014 nos va a permitir analizar el impacto de las BM-MSC isogénicas y alogénicas, así como sus VE en el modelo de trasplante renal en rata.
Limitaciones del uso de las vesículas extracelulares en la clínicaToda terapia basada en VE humanas se considerará medicamento biológico por contener una o más sustancias activas hechas o derivadas de una célula viva, y se someterá a las regulaciones de Europa, de Estados Unidos de América, Australia y Japón. Estas VE seguirán los estándares de seguridad para tejidos y células por tener en común con la fuente de origen su complejidad, composición y acción biológica. Sin embargo, teniendo en cuenta la relevancia de los resultados obtenidos a partir de los modelos animales, se asume que las terapias basadas en las VE no están incluidas en la definición de nuevos fármacos experimentales de elevado riesgo. Para la fabricación de dicha terapia es necesario contar con infraestructura, tecnología y un sistema de gestión de calidad para el cumplimiento de las normas GMP y GLP que tenga en cuenta la seguridad del donante y del receptor. Durante los estudios clínicos en fase inicial se monitorizarán la seguridad, la toxicidad y la inmunogenicidad de dicha terapia. Y será en estudios clínicos más avanzados (fase II-IV) en los que se estudiará la eficacia y los efectos adversos de las VE autógenas o alogénicas que darán soporte a la traslación de dicha terapia a la clínica95.
Si bien el uso de las VE producidas por células progenitoras o MSC está logrando un éxito nada despreciable en modelos preclínicos agudos, todavía hay algunas preguntas sin respuesta que tendrán que ser abordadas antes de introducir terapias derivadas de estas observaciones en el ámbito hospitalario. ¿Qué componente o cóctel de compuestos de las VE es responsable del efecto regenerativo? ¿Cómo tiene lugar la biodistribución de las VE y su tropismo hacia las células tubulares epiteliales? En cuanto a la producción, la reducida secreción por parte de las células productoras, la compleja caracterización del contenido y la presentación de antígenos HLA en superficie son sus principales escollos. El reconocimiento de alo-HLA puede ser un gran inconveniente de la terapia con VE, pero se están buscando estrategias para generar VE sin HLA o bien producirlas de forma sintética. La producción sintética de VE con un contenido ajustado a las necesidades (miRNA, mRNA, proteínas) que no sean reconocidas por el sistema inmune del paciente será el objetivo final.
Desarrollo de vesículas sintéticas para su uso clínicoLa producción de VE artificiales o sintéticas pasa por el uso de liposomas, micropartículas o nanopartículas compuestas por polímeros biodegradables como el ácido poli(láctico-co-glicólico) (PLGA)96–98, o colágeno99, itoh100 o dextrano101–103. Las vesículas sintéticas nos permitirán controlar mejor la liberación de los compuestos y el direccionamiento hacia los tejidos dañados, reducir los efectos secundarios e incrementar su biodisponibilidad para finalmente aumentar la calidad de vida del paciente104. Concretamente, los liposomas son el sistema más aceptado por su biocompatibilidad y biodegradación, gran solubilidad, elevada vida media, liberación selectiva en el lugar de acción y capacidad para resistir la acción de agentes quimioterapéuticos. Sin embargo, hasta el momento tienen baja especificidad, si bien hay estudios en los que se desarrollan liposomas sensibles a la temperatura, pH, luz, campo eléctrico o campo magnético, así como el acoplamiento de ligandos y anticuerpos a su membrana105.
La producción a gran escala de liposomas y su formulación para la clínica también presenta varios inconvenientes: la inestabilidad, toxicidad después de administraciones repetitivas y activación del complemento106,107. A pesar de estos inconvenientes, se han desarrollado varios sistemas de liberación de fármacos basados en liposomas que están en etapa preclínica, ensayos clínicos y algunos aprobados como tratamiento en clínica, concretamente en el campo de la quimioterapia108,109. A pesar de todos estos avances, el diseño de nuevas formulaciones liposómicas requiere estudios profundos tanto in vitro como in vivo para llevar a cabo los estudios preclínicos antes de su transferencia a la clínica.
Actualmente, nuestro grupo, junto a 3 centros europeos, está inmerso en el proyecto EV Stem Injury financiado a través del programa «FP7-PEOPLE-2013-IAPP-Marie Curie Action: Industry-Academia Partnerships and Pathways». Este proyecto incluye 2 enfoques diferentes para la producción de VE: biológicas y sintéticas. Con el fin de explorar el efecto renoprotector de las VE, se evaluará su potencia y eficacia en modelos in vitro e in vivo de la lesión renal aguda y crónica.
Conceptos clave- •
Las vesículas extracelulares son un sistema de comunicación entre células que comporta la transferencia de material proteico y genético.
- •
Existen distintos tipos de vesículas extracelulares en función de su origen y tamaño.
- •
Las células madre progenitoras y sus vesículas extracelulares presentan el mismo potencial terapéutico.
- •
El uso de vesículas extracelulares evitaría el riesgo de la mala diferenciación que supone el uso de células madre indiferenciadas.
- •
El potencial renoprotector de las vesículas extracelulares ha sido extensamente estudiado en modelos agudos, pero es necesario profundizar en su aplicación en modelos crónicos y de trasplante renal.
- •
El estudio del contenido de las vesículas extracelulares es imprescindible para concretar el componente o cóctel de componentes con efecto renoprotector.
- •
La fabricación de vesículas sintéticas con una composición más controlada y una capacidad terapéutica equivalente a las biológicas hará posible su futura aplicación en la práctica clínica diaria.
Los autores de este trabajo declaran la no existencia de conflicto de interés.
Los investigadores del LENIT forman parte de la REDinREN (RD12/0021/0028) del Instituto de Salud Carlos III-Ministerio de Ciencia e Innovación, cofinanciada por el Fondo Europeo de Desarrollo Regional (FEDER) «Una manera de hacer Europa».
JMC recibió una ayuda a la investigación de la S.E.N. en 2014.
Este trabajo se ha desarrollado en el Centre de Recerca Biomèdica Cellex, Barcelona, España.



