



Los microARN son pequeñas moléculas endógenas de ARN de vital importancia para la regulación de la expresión génica. Se han convertido en los mediadores biológicos más importantes que se han caracterizado en los últimos diez años. Participan en casi todos los procesos celulares, por lo que su desregulación está asociada al desarrollo de muchas patologías, entre las que se encuentran las renales. Existen cada vez más pruebas que demuestran que los microARN son reguladores claves de la función y el desarrollo renal, aunque también se encuentran en el origen de algunas enfermedades renales. Los estudios más recientes han concluido que estas moléculas pueden ser secretadas al exterior de la célula, lo que permite que puedan ser detectadas en fluidos periféricos como la orina y el suero. Además, los microARN circulantes detectados en los fluidos corporales pueden ser biomarcadores adecuados de las enfermedades renales, entre las que se incluye la lesión renal aguda. Esta nueva generación de biomarcadores renales podría tener consecuencias importantes para la práctica clínica, ya que podrían contribuir significativamente a la mejora del manejo de los pacientes. En este trabajo se revisa la implicación de los microARN en la homeostasis y la función renal y el papel de los microARN circulantes como nuevos biomarcadores de las enfermedades renales, centrándonos en su potencial utilidad para el manejo de la lesión renal aguda.
microRNAs are small, endogenous RNA molecules which are critical for a new step in the regulation of the gene expression. They have become the most critical biological mediators characterized in the last ten years. microRNAs participate in almost every cellular process, therefore their deregulation is associated with the development of a wide range of pathologies, including kidney diseases. Increasing evidence demonstrates that microRNAs are key regulators of the normal kidney function and development, but they are also at the basis of several renal diseases. Recent works have established that these molecules can be secreted to extracellular environments, enabling their detection in peripheral body fluids such as urine and serum. Moreover, circulating miRNAs detected in body fluids turn into suitable biomarkers of kidney diseases, including acute kidney injury. This new generation of renal biomarkers could have a great impact in the clinical practice, significantly contributing to improve patient management. In this review, we discuss over the implication of microRNAs in normal kidney function and homeostasis as well as the role of circulating miRNAs as novel biomarkers of kidney diseases, focusing on their potential usefulness in acute kidney injury management.
microRNAs
miRNAs are small (20-25 nucleotides) non-coding RNAs that have radically changed our understanding of gene post-transcriptional regulation. They are fine tuners of gene expression and more than 90% of the genes in mammals are submitted to their control, participating in almost every cellular function. Moreover, their deregulation is frequently associated with disease development.1
More than 2000 miRNAs have been described in the human genome,2 a number comparable to the known number of transcription factors or other regulatory proteins. Moreover miRNAs show very specific expression patterns among tissues and cell types.
Their mechanism of action is based on recognition of small sequences in their target mRNAs. This feature makes it possible for one miRNA to regulate hundreds of target mRNAs and that one mRNA can be regulated by several miRNAs. This dynamic regulation has unveiled them as key regulators of a wide range of cellular events, including rapid responses to stress.1
microRNA Biogenesis
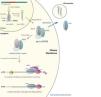
miRNAs are mainly transcribed by RNA polymerase II as longer primary transcripts called pri-miRNAs. miRNAs genes are often located in non coding DNA regions and they are frequently organized in clusters. Clustered miRNAs are transcribed as a single, longer pri-miRNA which generates several functional miRNAs by subsequent processing. miRNAs genes can also be found in protein coding genes, specially located inside introns. In these cases, splicing of the coding mRNAs leads to generation of the functional miRNA. As other transcripts produced by RNApolymerase II, pri-miRNAs present 5´Cap and 3´Poly-A Tail.
Pri-miRNAs molecules form imperfect stem-loop structures that can be recognized by a processing complex formed by the RNAse III enzyme Drosha and the RNA binding protein (RBP) DGCR8. Stem-loop double stranded structures of pri-miRNAs are recognized by DGCR8, which guides the positioning of Drosha. This catalytic centre cleaves pri-miRNAs liberating a hairpin RNA molecule of 70-100 nucleotides known as pre-miRNA.
Pre-miRNAs are exported to the cytoplasm by the nuclear export receptor Exportin 5 in a Ran-GTP dependent manner. Once in the cytoplasm pre-miRNAs are further processed by another RNAse III enzyme called Dicer. A new cleavage produces a double stranded RNA molecule of 22 nucleotides. One of the strands (the mature miRNA) is transferred to the Argonaute protein to conform the RNA Induced Silencing Complex (RISC). The other strand (often called minor, passenger or *strand) is frequently degraded. Strand selection mechanism has not been completely elucidated yet. However, it has been proposed that the strand with the less stable base-pairing in its 5´end is often chosen as guide strand to be loaded in the RISC complex.
RISC complex is the key effector of miRNA regulation. This complex, joined to other controlling factors, is responsible for driving mRNA degradation or translation repression, as will be further detailed in following sections.1,3
miRNA Target Recognition and Function
Once loaded into the RISC complex, miRNAs recognize their target mRNAs by base-pair complementarity. Target sequences are mainly located in the 3´ untranslated region (UTR) of mRNAs. However, functional miRNA binding sites can also be found in the 5´ UTR and open reading frame regions.
Nucleotides in positions 2-8, called seed sequence, are essential for pairing with the target mRNA and miRNA function. In the case of perfect complementarity of the seed sequence of the miRNA with the target sequence, miRNAs act as a short interfering RNA (siRNA) promoting mRNA cleaving by RISC complex. This mechanism is principally found in plants but is very rare in mammals.
When pairing with target sequences is partially complementary, which is the most frequent mechanism in mammals, miRNA regulation is produced by mRNA translation repression or degradation. However this degradation process is different and involves recruitment of deadenylase complexes that remove or shorten the poly-A tail of the target transcript. Poly-A tail shortening induces decapping of the 5´ extreme of the transcript and uncapped mRNAs are rapidly degraded by 5´ to 3´ exoribonucleases (Figure 1).1,3
Although the contribution rate of mRNA decay and translational repression to miRNA action is a controversial topic, it seems clear that target degradation provides a major contribution to silencing in mammal cells. In this regard, it has been estimated that mRNA decay is present in 85% of the miRNA regulation process whereas 15% corresponds to translational repression.3,4
miRNA Decay and Turnover
Accumulative evidence reveals that different miRNAs present distinct and inherent half-lives that could be encoded by their sequence or can be determined after maturation by post-transcriptional mechanisms such as uracile and adenosine addition to their 3´end.
Half-lives of some miRNAs could reach many hours or even days in some organs like the liver or the heart.5,6 However, their ability to regulate rapid cell responses to environmental signals makes this slow turnover rate not appropriate for every context. Indeed, some miRNAs expressed in the retina and involved in darkness adaptation present a half-life of approximately 1 hour.7
These interesting data point out that more attention needs to be focused on miRNA turnover as miRNA half-life regulation could emerge as a potential critical step in miRNA function. Indeed, miRNA decay regulation is one of the big challenges in miRNA biology for the next years.
microRNAs IN KIDNEY
Due to the relevance of miRNA function and implication in a wide range of processes such as organ development, homeostasis and pathophysiology, miRNA studies in kidney development and function have emerged as a field of intense research during the last few years.
microRNAs in Normal Kidney Function and Development
Several profiling studies have revealed microRNA profiles related to specific organs, including the kidney. These studies indicate that miR-146a, miR-886, miR-192, miR-194, miR-204, miR-215 and miR-216 are renal-specific microRNAs. In addition, let-7a-g, miR-196a/b, miR-10a/b, miR-130, miR146, miR-21, miR-200a, miR-30a-e and miR-872 are highly expressed in renal tissue.8-11
microRNAs are essential for kidney development. In fact, lack of miRNA activity causes defects in kidney terminal differentiation such as decreased proliferation rate, aberrant nephron patterning and delayed terminal differentiation of kidney tubules. In this regard, miR-30 and miR-200 family strictly regulate the temporal and spatial expression pattern of transcription factors involved in pro-nephron maturation.12
Loss of microRNAs in nephron progenitors leads to early depletion of this cell population as renal growth progresses. These effects are especially due to the regulation by miR-10a, miR-106b and miR-17-5p of Bim, a pro-apoptotic factor which increases its expression levels when miRNAs are depleted.13
On the other hand, some microRNAs have been related to kidney senescence. Particularly, overexpression of miR-335 and miR-34a in mesangial cells leads to premature senescence of this cell population by the regulation of mitochondrial antioxidative enzymes.14
Studies based on DICER knockdown in podocytes, juxtaglomerular cells and proximal tubules have unveiled the role of miRNAs in kidney function maintenance as well as specific miRNA roles in each kidney compartment. KO of DICER in glomerulus results in progressive loss of podocyte function leading to proteinuria and kidney function impairment and ultimately leading to animal death. Three microRNAs in particular have been identified as contributors to this renal failure (miR-23b, miR-24 and miR-26a).13,15 In addition, Drosha ablation in podocytes results in proteinuric renal disease and collapsing glomerulopathy.16
DICER deletion in juxtaglomerular cells produces acute loss of this cell type leading to an abrupt decrease of renin expression in kidney and deregulation of blood pressure.17
Several microRNAs have also been unveiled as critical regulators of kidney function and homeostasis maintenance. For instance, miR-320a is a direct regulator of Aquoporin 1 and 4, which are integral membrane transporters involved in water homeostasis.18 Moreover, the Na/H exchange factor-1, a regulator of ion transport in apical membrane, is a target gene of miR-200b.19
All these studies indicate that miRNAs play critical roles in normal renal function and physiology maintenance and when altered, may lead to renal diseases.
microRNAs in Kidney Disease
Specific miRNAs deregulation has been linked to renal disease development. For example, miR-192 has been identified as a critical regulator of collagen production in diabetic nephropathy.20 Studies in renal biopsies from diabetic patients have shown that miR-192 is poorly expressed in renal tissue. This downregulation is directly correlated with tubulointerstitial fibrosis and low glomerular filtration rate (GFR) in diabetic patients.21
Several microRNAs have been related to polycystic kidney disease (PCK). miR-10a, miR30a-5p, miR-96, miR-126-5p, miR-182, miR-200a, miR204, miR-429 and miR-488 were downregulated in a Pkd1 -/- mouse model.22 Moreover, miR-15 expression was decreased in a rat model of PCK. Reduction of this microRNA increases the expression of Cdc25A, a regulator of cell cycle, promoting cyst growth.23
microRNA deregulation has also been associated with renal carcinoma. miRNA profiling experiments demonstrated that miR-28, miR-185, miR-7-2 and let-7f-224 and miR-155, miR-210, miR-592, miR-34b, miR-224, miR142-3p, miR-185, miR-34a, and miR-21 were modulated in renal carcinoma tissue samples.25 All these microRNAs show target genes enriched in angiogenesis, hypoxia, apoptosis, tumorogenesis, invasion and metastasis pathways, in agreement with the pathological context where they have described.26
On the other hand, the miR-200 family are repressors of E-cadherin during epithelial to mesenchimal transition (EMT), promoting cell migration and invasion during metastasis.27,28 In addition, miR-200a and miR-141 are involved in EMT and fibrosis mediated by TGFb1 in vivo and in vitro, and have an important implication in renal fibrosis development as demonstrated with animal experimental models.29 These miRNAs could be considered as novel therapeutic target in EMT-associated nephropathies including chronic kidney diseases, where efficient treatments are still not available.
miRNAs in Renal Ischemia/Reperfusion
The clear role of miRNAs in kidney function and nephropathies and the ability of miRNAs to regulate rapid responses to stress and injury have unveiled microRNAs as potential key regulators of kidney responses to acute damage, such as Acute Kidney Injury following renal Ischemia/Reperfusion (I/R).
miRNAs involvement in kidney response to I/R was undoubtedly demonstrated by DICER KO experiments in proximal tubular cells. DICER KO mice showed normal renal function and proximal tubule structure in basal conditions. However, general downregulation of miRNAs conferred a protection against I/R injury, since DICER KO animals subjected to I/R exhibited significantly better renal function, less tissue damage, lower tubular apoptosis and improved survival compared to wild-type animals.30
Global expression profiling experiments in mice undergoing renal I/R have identified a signature of 9 microRNAs (miR-21, miR-20a, miR-146a, miR-199a-3p, miR-214, miR-192, miR-187, miR-805, and miR-194) which are differentially expressed in kidney compared to sham controls.31 Moreover, a more recent work from the same group demonstrates that microRNA signature after I/R is different in operated animals when compared to sham controls and these differences are maintained and evolve along time. Their data strongly evidence that altered miRNAs after I/R could be considered as new biomarkers for renal I/R injury.32
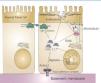
In addition to the miRNAs signature of renal response to I/R, miRNAs also have been implicated in protection against ischemic injury, showing that these molecules are potential novel therapeutic targets in this context. In fact, our lab recently demonstrated that miR-127 is a regulator of the proximal tubule cell response to I/R injury. Overexpression of this microRNA in an in vitro model of I/R demonstrates that miR-127 prevents focal adhesion complex disassembly and thigh junction disruption provoked by I/R. Moreover, kinesin familiy member 3B, a component of the Kinesin II complex involved in intracellular vesicle trafficking, is a target gene of miR-127 in proximal tubule cells during I/R, with important implications in kidney function.33 Based on these results, the potential role of miR-127 in the pathophysiology of ischemic proximal tubule injury is summarized in Figure 2.
Moreover, Liu F. et al. demonstrate that miR-210, a well established miRNA regulated by hypoxia, is involved in angiogenesis regulation during renal I/R by activating VEGF and its receptor VEGFR2 expression, contributing to ischemic injury recovery.34 Regarding to vascular compartment involvement in renal I/R response, miRNAs contained in vesicles derived from endothelial progenitors protect renal cells from I/R injury. Intravenous injection of microvesicles confers functional and morphologic protection of renal cells by enhancing tubular cell proliferation and reducing apoptosis and leukocyte infiltration. These miRNAs also protect kidney from chronic damage progression by inhibiting glomerulosclerosis and tubulointerstitial fibrosis. Thus, vesicle-derived miRNAs contribute to reprogram resident renal cells to a regenerative program after I/R injury.35
CIRCULATING microRNAs AS NOVEL BIOMARKERS
Even though the initial studies proposed that miRNAs were only present inside the cell, it has been demonstrated that they can also be secreted to the extracellular environment with potential functional consequences.36 This secretion process has resulted in their detection in a wide range of cell-free body fluids such as breast milk, urine or serum.37
miRNA secretion seems a highly regulated process and selection of miRNAs which can be secreted is not a random process. On the other hand, as mentioned before, miRNA deregulation has been associated with the development of a wide range of pathologies. Due to these features, physiological or pathological regulation of intracellular miRNAs may also modify the panel of secreted miRNAs. Indeed, changes in serum miRNAs profiles have been unveiled as useful markers of a wide range of diseases including cancer, cardiovascular disease, stroke and nephropathies as well as physiological states such as pregnancy.38
Moreover, serum miRNAs have demonstrated great stability and resistance to aggressive conditions such as RNAse treatment, or drastic pH changes.39 The mechanism underlying this unexpected miRNA stability in serum is still not completely understood, but it has been evidenced that circulating microRNAs are released from cells in membrane vesicles which protects them from the environment. Vesicles proposed to be carriers of circulating microRNAs include exosomes, which are 50-90nm vesicles released by exocytosis from multivesicular bodies (MVB)36,40 as well as larger microvesicles up to 1µm.41,42 Furthermore, recent studies have demonstrated that serum circulating miRNAs can also be carried by Argonaute2 proteins.43
Taken together, all these features demonstrate that serum circulating miRNAs achieve nearly all the required characteristics for an ideal biomarker. Their presence in a peripheral fluid allows diagnosis by minimum invasive methods and samples can be easily and routinely obtained in clinical practice. In addition to their high stability in fresh samples, several studies have demonstrated that miRNAs maintain stability and reliability in long-term stored serum samples and in samples conserved at room temperature for hours.39 Moreover, they can be easily quantified by qRT-PCR, with affordable technical effort. These characteristics, joined to their tissue and cell type specificity, have revealed circulating miRNAs as promising biomarkers for more accurate diagnosis and monitoring of diseases.
Regarding the advantage of circulating miRNAs as precise biomarkers in comparison to the ones available up to now, it is important to notice that miRNAs could belong to the pathophysiological mechanisms responsible for diseases, including renal diseases. Therefore, miRNAs could provide the clinician with critical additional information that is currently not accessible and they could also be considered as novel therapeutic targets.
CIRCULATING microRNAs AS AKI BIOMARKERS
Despite the intense research in the circulating miRNAs field of the last years, publications about their role as AKI biomarkers are still scarce. However, during the last two years some articles have started to point out this issue.
Lorenzen JM et al. in 2011 revealed that miR-210 was upregulated in plasma samples of a cohort of 77 AKI patients compared to healthy controls. Moreover, plasma levels of this microRNA could predict patient survival 4 weeks after initiation of Renal Replacement Therapy (RRT).44
In addition, miR-21 and miR-155 are upregulated in renal tissue after I/R or toxic injury, correlating with a lower expression in blood and urinary supernatants. Moreover, these microRNAs were also modulated in urine samples of AKI patients compared to healthy volunteers. Bioinformatics analysis of the target genes of these microRNAs evidenced that they were enriched in pathways related to apoptosis and cell proliferation.45
A recent publication by Lan YF et al., demonstrates that miR-494 is a regulator of the renal inflammatory response as well as apoptosis after I/R injury. Moreover, detection of this miRNA in urine showed that miR-494 expression is 60 fold higher in AKI patients compared to healthy controls.46
In addition to these reports, our group has identified and validated a panel of 10 serum miRNAs, including miR-101-1, miR-127-3p, miR-210, miR-126, miR-26b, miR-29a, miR-146a, miR-27a, miR-93* and miR-10a, as biomarkers of AKI, using cohorts of intensive care units (ICU) patients with AKI and cardiac surgery with cardiopulmonary bypass patients. ROC analysis for diagnostic value of the panel demonstrated sensitivity and specificity close to 100%. Moreover, serum miRNAs detect AKI development before serum creatinine increases, becoming early diagnostic tools. These miRNAs provide additional valuable clinical information such as AKI severity, AKI origin (pre-renal or intrinsic) or AKI aetiology. Moreover, serum miRNAs could be considered biomarkers of AKI predisposition since they can predict AKI development when when microRNA are estimated before cardiac surgery.47
All these studies demonstrated that miRNAs could be precise biomarkers of AKI diagnosis useful in clinical practice, offering valuable additional information in comparison with classical biomarkers. Since miRNAs are capable of indicating survival, recovery, AKI origins or AKI aetiologies, they could significantly contribute to improve AKI patient management.
FUTURE PERSPECTIVES OF miRNAs IN AKI
The information provided here identifies miRNAs as very promising molecules for increasing the knowledge of pathophysiological mechanisms underlying kidney diseases and very powerful tools for improving clinical management of renal patients.
miRNAs identification and validation as biomarkers for nephropathies will contribute to more precise diagnostics and prognostics as well as will provide valuable additional information for patient handling. Although it was not the main topic of this review, miRNAs could also be considered as novel therapeutic targets in renal diseases. In this regard, the improved knowledge of miRNAs involved in renal I/R injury and recovery could be a key point for developing new therapeutic approaches to prevent AKI, reduce chronic renal disease derived from acute episodes and improve outcome.
Identification of microRNAs as novel biomarkers for AKI could drastically change current clinical practice allowing earlier detection and better patient monitoring and handling. Therefore, taking into account the reported results and our findings, the use of miRNAs in AKI clinical practice can be proposed, based on the dynamic model of AKI by Pickering and Endre48 (Figure 3). This model establishes the concept of Phase-Specific AKI Biomarkers.
miRNAs could be considered Phase-1 Biomarkers since they could identify patients in risk of AKI development,47 thus allowing the application of preventive strategies. miRNAs could also be Phase 2 Biomarkers since miRNAs expression correlates with AKI diagnosis before creatinine increases, permitting the application of early therapeutic interventions. Phase 3 Specific Biomarkers are able to monitor injury and repair. In this regard, serum miRNA levels correlate with severity and different recovery degrees.45,47 Finally, Phase 4 Specific Biomarkers, including serum miRNAs, will identify AKI outcomes such as CKD development, RRT requirement or survival.44-47
In summary, miRNAs have started to emerge as key players in many relevant diseases, including nephropathies. This review has aimed to introduce and overview miRNAs as exciting molecules in Experimental Nephrology and promising tools in Clinical Nephrology.
KEY CONCEPTS
- microRNAs are small RNA molecules which are key regulators of gene expression.
- microRNAs are involved in normal kidney function and development as well as in development of renal pathologies.
- microRNAs can be detected in peripheral body fluids such as serum or urine.
- Circulating microRNAs detected in body fluids are novel biomarkers of renal diseases, including Acute Kidney Injury.
FUNDING
This work was supported by grants FIS PS09/02183 and PS12/00094 by Instituto de Salud Carlos III (ISCIII) and Ayuda Intramural FIBIO HRYC 122/2009. EAF is supported by CAM-CIFRA (S2010/BMD-2378), ER is funded by REDINREN (RD12/0021/0020), MR is recipient of contract from ISCIII (CA11/00491). We are also grateful to F. Jiménez for his valuable help with figures editing.
Conflicts of interest
The authors declare that they have no conflicts of interest related to the contents of this article.

Figure 1. microRNA biogenesis, function and secretion.

Figure 2. miR-127 as potential mediator of AKI pathophysiology in proximal tubule cells.

Figure 3. Circulating miRNAs as useful biomarkers of AKI in clinical practice.



