

The intestinal microflora maintains a symbiotic relationship with the host under normal conditions, but its imbalance has recently been associated with several diseases.
In chronic kidney disease (CKD), dysbiotic intestinal microflora has been reported with an increase in pathogenic flora compared to symbiotic flora. An enhanced permeability of the intestinal barrier, allowing the passage of endotoxins and other bacterial products to the blood, has also been shown in CKD. By fermenting undigested products that reach the colon, the intestinal microflora produce indoles, phenols and amines, among others, that are absorbed by the host, accumulate in CKD and have harmful effects on the body. These gut-derived uraemic toxins and the increased permeability of the intestinal barrier in CKD have been associated with increased inflammation and oxidative stress and have been involved in various CKD-related complications, including cardiovascular disease, anaemia, mineral metabolism disorders or the progression of CKD. The use of prebiotics, probiotics or synbiotics, among other approaches, could improve the dysbiosis and/or the increased permeability of the intestinal barrier in CKD.
This article describes the situation of the intestinal microflora in CKD, the alteration of the intestinal barrier and its clinical consequences, the harmful effects of intestinal flora-derived uraemic toxins, and possible therapeutic options to improve this dysbiosis and reduce CKD-related complications.
La microflora intestinal mantiene una relación simbiótica con el huésped en condiciones normales, sin embargo, su alteración se ha asociado recientemente con numerosas enfermedades.
En la enfermedad renal crónica (ERC) se ha descrito una disbiosis en la microflora intestinal con un aumento de la flora patógena sobre la simbionte. Además, la permeabilidad de la barrera intestinal está aumentada, lo que permite el paso de endotoxinas y otros productos bacterianos a la sangre. La microflora intestinal, mediante la fermentación de productos no digeridos que alcanzan el colon, produce indoles, fenoles, o aminas, entre otros, que son absorbidos por el huésped, se acumulan en la ERC y tienen efectos deletéreos sobre el organismo. Estas toxinas urémicas generadas en el intestino y el aumento de la permeabilidad de la barrera intestinal en la ERC se han asociado a un aumento de la inflamación y el estrés oxidativo, y están implicados en diversas complicaciones asociadas a la ERC, como la enfermedad cardiovascular, la anemia, las alteraciones del metabolismo mineral o la progresión de la ERC. El uso de prebióticos, probióticos o simbióticos, entre otras aproximaciones, podrían mejorar la disbiosis o el aumento de la permeabilidad de la barrera intestinal en la ERC.
En este artículo se revisan la situación de la microflora intestinal en la ERC, la alteración de la barrera intestinal y sus consecuencias clínicas, los efectos deletéreos de las toxinas urémicas derivadas de la microflora intestinal, así como las posibles opciones terapéuticas para mejorar esta disbiosis y reducir las complicaciones de la ERC.
Since Hippocrates (400 BC) established that “death lies in the intestines”, their influence on the health of human beings has been well known.
The germs that inhabit our body are called the microbiota, and their collective genomes, the microbiome. More than 100 trillion germs (1014) cohabit with us throughout our lives, representing 10 times the number of cells that make up our body and constituting 1.5–2kg of our weight.1,2 The concentration of germs in the digestive tract gradually increases from the stomach to the colon, where they reach the highest concentration (up to 1011 microorganisms/g of faeces) and diversity. The gut microbiota plays an important role in metabolic, nutritional, physiological, and immunological processes, and constitutes a true ecosystem.3 The human microbiome is our second genome, which has more than 3 million genes (100 times more genes than the human genome itself) and is the subject of research by the Human Microbiome Project Consortium.4–6
Originally, the gut microbiota is formed through the placenta, where low levels of non-pathogenic germs, especially the phyla firmicutes, bacteroidetes, and Fusobacteria are nested. In the first years of life, feeding, type of birth, hygiene, and use of antibiotics condition the formation of the intestinal microbiome.7,8 Different species of germs colonise and are originated during various events (Table 1).
Intestinal microflora in relation to perinatal events.4,5
| Exposure | Intestinal flora |
|---|---|
| Vaginal canal | Bifidobacterium, bacteroids, Lactobacillus, prevotella, enterococci, streptococci, Clostridiaeceae |
| Post-caesarean delivery | Staphylococcus, Corynebacterium Propionibacterium. Lower amount of Bifidobacterium and bacteroides |
| Breast-feeding | Bifidobacterium, bacteroides, Lactobacillus, clostridia, actinobacteria and firmicutes |
| Artificial breastfeeding | Bacteroides, clostridia, Enterobacteriaceae |
The gut microbiota is established in the first 2–3 years of life as a dynamic ecosystem, dominated by bifidobacteria; their composition increases in richness and diversity until reaching their maximum complexity in adulthood, when the dominant species are bacteroidetes, firmicutes and actinobacteria.9–11
Bacterial communities that lie in the intestine are, therefore, a combination of different types and amounts of bacteria and 3 different groups of microbiota or enterotypes have been identified in humans.12 The phylogenetic composition of intestinal microflora tends to be similar between individuals from the same region, from the same family and also with a similar diet which plays a significant role in their composition.13,14
Dietary habits affect the composition of the gut microbiota. Since the microbiota is in contact with a significant number of neural cells and immunological cells, it directs the maturation of the immune system in childhood and contributes to the maintenance of its homeostasis during life.2 Complex polysaccharides, which are not digested by enzymes in the small intestine, are metabolised by the colon microflora. These polysaccharides are degraded and fermented in the large intestine and converted into short chain fatty acids (SCFAs) and gases (CO2 and H2). A high intestinal content of fructose promotes the formation of butyrate which is produced by bacteria. Dietary supplementation with specific polysaccharides may promote the growth of “healthy” germs (Bifidobacterium, Lactobacillus), the production of SCFA and it may decrease intestinal pH that inhibits the growth of pathogenic bacteria.15,16
In the ageing process, progressive damage occurs to the morphology and function of the different systems and the microbiota becomes less diverse and more dynamic, characterised by the predominance of bacteroidetes over firmicutes, with an increase of Protobacteria spp. and a decrease of Bifidobacterium. This is evidenced by rRNA techniques and it has been analysed as part of the study ELDERMET.17–19 Major changes in the microbiota are found in the colon of persons over 60 years of age. The significance of these changes is yet to be clarified.20
Faecal bacteria such as Escherichia coli, which divides every 20min, are genetically prepared to be highly adaptative, and always survive even though their host ages. Without this plasticity, we probably would not have been able to cope with changes in lifestyle and dietary habits, as evidenced by the transition from the Palaeolithic to the dietary habits of Western societies.21
Functionally, the gut microbiota provides nutrients and energy to the body through fermentation of nondigestible foods in the large intestine. The most important fermentation products deriving from the fermentation are the SCFA, which serve as a source of energy to intestinal cells and bacteria, and contribute to energy expenditure, satiety, and glucose homeostasis.22 Other relevant functions of the gut microbiota are the endogenous synthesis of certain vitamins and amino acids, the metabolism of bile acids, or the maintenance of the integrity of intestinal barriers, which protect the host from pathogenic germs.
Thus, the gut microbiota is involved in the maturation of the immune system in infancy and contributes to the maintenance of its homeostasis throughout life.23
Gut microbiota in chronic kidney diseaseFrom the early stages of chronic kidney disease (CKD) there is a quantitative and qualitative alteration of intestinal microflora (dysbiosis); so the composition and metabolic activities of microflora are changed in CKD and this is a hot and innovative topic in nephrology literature. These alterations include changes in intestinal transit, decreased protein absorption, decrease in dietary fibre intake, treatment with oral iron and frequent use of antibiotics.
All of this contributes to systemic inflammation and the accumulation of uraemic toxins that are absorbed by intestine and eliminated by the kidney. Inflammation and uraemic toxins play a central role in the pathophysiology of atherosclerosis, as well as in other complications associated with CKD24–27; that will be reviewed below.
Patients with CKD are polymedicated. Some drugs frequently prescribed to these patients may alter intestinal microflora, especially antibiotics,28,29 but others may also slow intestinal transit, phosphorus binders, ion exchange resins,30 or iron supplements that may have an effect on microflora but it is not well defined.31,32
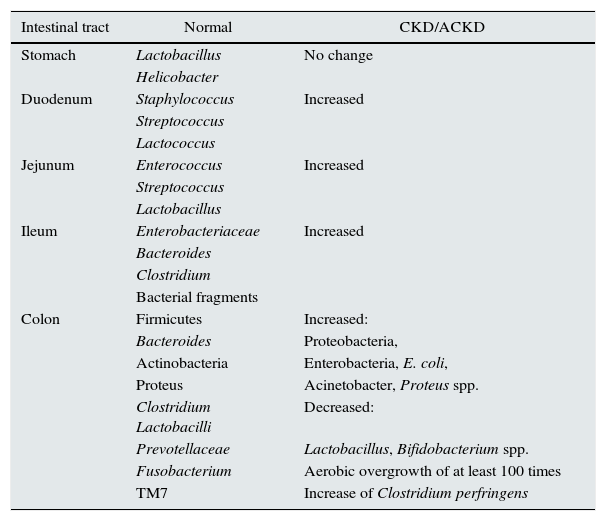
Change in intestinal barriers in chronic kidney diseaseChanges in intestinal barriers with an increased intestinal permeability is common in CKD (Table 2).
Gut microbiota changes in CKD.
| Intestinal tract | Normal | CKD/ACKD |
|---|---|---|
| Stomach | Lactobacillus | No change |
| Helicobacter | ||
| Duodenum | Staphylococcus | Increased |
| Streptococcus | ||
| Lactococcus | ||
| Jejunum | Enterococcus | Increased |
| Streptococcus | ||
| Lactobacillus | ||
| Ileum | Enterobacteriaceae | Increased |
| Bacteroides | ||
| Clostridium | ||
| Bacterial fragments | ||
| Colon | Firmicutes | Increased: |
| Bacteroides | Proteobacteria, | |
| Actinobacteria | Enterobacteria, E. coli, | |
| Proteus | Acinetobacter, Proteus spp. | |
| Clostridium Lactobacilli | Decreased: | |
| Prevotellaceae | Lactobacillus, Bifidobacterium spp. | |
| Fusobacterium | Aerobic overgrowth of at least 100 times | |
| TM7 | Increase of Clostridium perfringens |
CKD, chronic kidney disease; ACKD, advanced chronic kidney disease.
Increasing urea levels and expanding bacteria with urease causes an increase ammonium production in the intestinal lumen and induce changes in intestinal pH that produces an alteration of intestinal permeability by affecting the tight junctions of the enterocyte. Vaziri et al. have demonstrated a marked reduction of tight junction proteins, claudin-1, occludin, and ZO1, in the colonic mucosa in CKD; this is associated with an infiltration of mononuclear leukocytes in the lamina propria and a marked thickening of the colon wall.33 There is histological evidence of chronic inflammation of the intestinal tract including oesophagitis, gastritis, etc.33,34
The presence of frequent oedema and hypervolaemia in CKD may aggravate intestinal barrier dysfunction in CKD patients on, haemodialysis, or peritoneal dialysis. In addition, excessive ultrafiltration and episodes of hypotension during haemodialysis may cause episodes of transient intestinal ischaemia which increases the permeability of the intestinal barrier facilitating the passage of endotoxins.33
In renal transplant patients, the investigation of the gut microbiota is in its infancy. It is known that inflammatory processes, such as graft ischaemia time, baseline disease and immunosuppressive drugs may play a relevant role in the alteration of the intestinal barrier.35,36
Intestinal microflora as a cause of inflammation in chronic kidney diseaseIn CKD, the decreased clearance of proinflammatory cytokines, is associated with the development of oxidative stress and inflammation. The later are contributing factors to the progression of the disease and its complications, including cardiovascular disease, cachexia, and anaemia. Oxidative stress and chronic inflammation stimulate the NF-κB transcription factor, which is the key regulator of proinflammatory cytokines and chemokines. Increased permeability of intestinal barriers in CKD patients favours the translocation of bacterial products of intestinal origin, as evidenced by the presence of DNA fragments of circulating intestinal pathogens (aerobic and anaerobic), both in patients in different stages of CKD and on renal replacement therapy.37–39 The increase in circulating bacterial products of intestinal origin activates innate immunity, promotes the inflammatory state associated with CKD and, increases the incidence of cardiovascular disease and mortality.40–42
Microbiota and uraemic toxins derived from the intestine in chronic kidney diseaseIntestinal production of uraemic toxinsThe origin of uraemic toxins in CKD is multiple. The importance of toxins generated by intestinal microbial metabolism43 is increasingly recognised. Approximately 10g of proteins reach the colon daily, where they are degraded by intestinal bacteria to metabolites such as ammonium, amines, thiols, phenols and indoles. These colon fermentation products are eliminated through faeces, although a portion is absorbed and eliminated by the kidney, so these are accumulated in CKD.44 In CKD, the uraemic toxins derived from intestinal microflora are:
Phenols and indoles: p-cresol and indoxyl sulfate. Phenols include p-cresol, p-cresyl sulfate (PCS), p-cresyl glucuronide, phenylacetic acid, phenyl sulfate and phenol.45
- -
p-Cresol/p-cresyl sulfate: products of phenylalanine and tyrosine metabolism of intestinal anaerobic bacteria. p-Cresol is conjugated in the intestinal wall to PCS and p-cresyl glucuronide in the liver. PCS is the main circulating metabolite of p-cresol.46
- -
Phenol: mainly derives from ingestion, from the catabolism of tyrosine by intestinal bacteria, as well as from tobacco consumption.
- -
Phenylacetic acid: is the result of the degradation of phenylalanine.
Among the indoles are indoxyl sulfate (IS) and indoleacetic acid.45 Both originate from the degradation of tryptophan by intestinal bacteria and are subsequently sulfated in the liver into IS. Indoles and phenols are uraemic toxins bound to proteins.47
Amines and polyamines: amines and polyamines are generated from intestinal microbial metabolism. An amino acid that is clinically relevant and of growing interest is trimethylamine N-oxide (TMAO). TMAO is produced by the intestinal metabolism of quaternary amines, such as choline/phosphatidylcholine, betaine, or l-carnitine. l-Carnitine, which is present in red meat, also induces the formation of TMAO and it is associated with an increase in cardiovascular disease.48 Dietary sources of TMAO are red meats, meats in general, egg yolks, liver, dairy products and saltwater fish. In CKD, TMAO accumulates and its levels depends on the glomerular filtration, but its binding to proteins is low, and it is well eliminated with dialysis.
Polyamines are organic cations including cadaverine, spermine, spermidine and putrescine. They come from the decarboxylation of l-arginine, l-ornithine or lysine in the intestine. In CKD patients, putrescine, spermidine, and spermine are increased in serum.49 These molecules have been shown to interact with insulin and lipoproteins, and contribute to the acceleration of atherosclerosis along with other factors such as hypertriglyceridemia.50
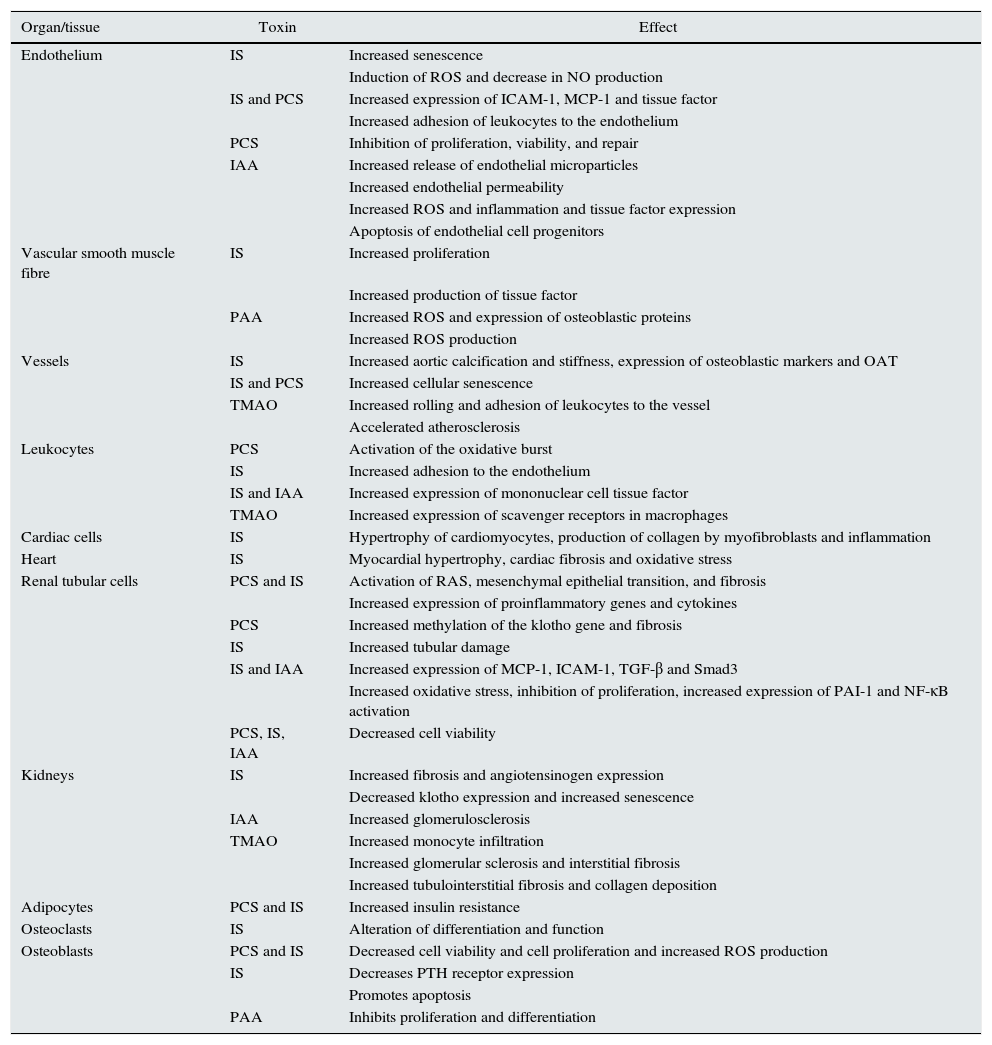
Biological and clinical consequences of the accumulation of uraemic toxinsThe aforementioned uraemic toxins have been associated with deleterious biological effects in different tissues and cell lines51,52 (Table 3), and with an increased risk of the progression of CKD, morbidity and mortality.
- a)
Progression of CKD: both IS and PCS are associated to the development of fibrosis, deterioration of renal function and disease progression.52,53 In vitro studies have shown a deleterious effects of these molecules on renal tubular cells.54 In a prospective study in patients with stage 1–5 CKD, the predictive role of both molecules in disease progression was confirmed.55 In experimental animals, a diet rich in choline or TMAO preduces progressive tubulointerstitial fibrosis and renal dysfunction.56
- b)
Cardiovascular complications: in CKD patients, IS is associated with endothelial damage, arterial stiffness and aortic calcification57; and, in hemodialysis patients it is associated with atherosclerosis58 and endothelial dysfunction,59 it has a cardiac profibrotic effect, favours hypertrophy of myocardiocytes60 and it is a predisposing factor of atrial fibrillation.61 Similar vascular effects have been described with,62 which is a predictor of cardiovascular risk in CKD patients.63,64 In hemodialysis patients PCS and IS have been associated with peripheral vascular disease and thrombosis of vascular access.65 A recent meta-analysis confirms the relationship of these molecules to cardiovascular risk in CKD.66
Likewise, indoleacetic acid is associated with oxidative stress and inflammation markers, and it is a predictor of mortality and cardiovascular events in CKD.67
Elevated levels of TMAO predicts coronary atherosclerotic burden56 and mortality in patients with CKD,68,69 although this is not shown in all reports.70
- c)
Anaemia: IS has been associated with anaemia of the renal patient; it interfere with the adequate production of erythropoietin71,72 and increased eryptosis (programmed cell death of red blood cells).73 Polyamines are associated to anaemia in renal patients, through an intra-erythrocytic effect,74 reduces erythropoiesis, and inhibit the activity of erythropoietin.
- d)
Alterations of bone-mineral metabolism: IS reduces bone formation by promoting oxidative stress in osteoblasts and inducing resistance to PTH, which favours the development adynamic bone.75 There is a positive correlation between FGF-23 and IS serum levels, suggesting an association between this molecule and metabolic bone disease in uraemic patients.76 Likewise, less bone remodelling has been observed in uraemic rats with higher IS after a parathyroidectomy.77
- e)
Insulin resistance: In CKD patients the catabolism of insulin is reduced and often, they also have insulin resistance, which is associated with an increased risk of mortality; it seems that insulin resistance is related to some of the uraemic toxins.78
Effects of different uraemic toxins at the cellular and tissue level.
| Organ/tissue | Toxin | Effect |
|---|---|---|
| Endothelium | IS | Increased senescence |
| Induction of ROS and decrease in NO production | ||
| IS and PCS | Increased expression of ICAM-1, MCP-1 and tissue factor | |
| Increased adhesion of leukocytes to the endothelium | ||
| PCS | Inhibition of proliferation, viability, and repair | |
| IAA | Increased release of endothelial microparticles | |
| Increased endothelial permeability | ||
| Increased ROS and inflammation and tissue factor expression | ||
| Apoptosis of endothelial cell progenitors | ||
| Vascular smooth muscle fibre | IS | Increased proliferation |
| Increased production of tissue factor | ||
| PAA | Increased ROS and expression of osteoblastic proteins | |
| Increased ROS production | ||
| Vessels | IS | Increased aortic calcification and stiffness, expression of osteoblastic markers and OAT |
| IS and PCS | Increased cellular senescence | |
| TMAO | Increased rolling and adhesion of leukocytes to the vessel | |
| Accelerated atherosclerosis | ||
| Leukocytes | PCS | Activation of the oxidative burst |
| IS | Increased adhesion to the endothelium | |
| IS and IAA | Increased expression of mononuclear cell tissue factor | |
| TMAO | Increased expression of scavenger receptors in macrophages | |
| Cardiac cells | IS | Hypertrophy of cardiomyocytes, production of collagen by myofibroblasts and inflammation |
| Heart | IS | Myocardial hypertrophy, cardiac fibrosis and oxidative stress |
| Renal tubular cells | PCS and IS | Activation of RAS, mesenchymal epithelial transition, and fibrosis |
| Increased expression of proinflammatory genes and cytokines | ||
| PCS | Increased methylation of the klotho gene and fibrosis | |
| IS | Increased tubular damage | |
| IS and IAA | Increased expression of MCP-1, ICAM-1, TGF-β and Smad3 | |
| Increased oxidative stress, inhibition of proliferation, increased expression of PAI-1 and NF-κB activation | ||
| PCS, IS, IAA | Decreased cell viability | |
| Kidneys | IS | Increased fibrosis and angiotensinogen expression |
| Decreased klotho expression and increased senescence | ||
| IAA | Increased glomerulosclerosis | |
| TMAO | Increased monocyte infiltration | |
| Increased glomerular sclerosis and interstitial fibrosis | ||
| Increased tubulointerstitial fibrosis and collagen deposition | ||
| Adipocytes | PCS and IS | Increased insulin resistance |
| Osteoclasts | IS | Alteration of differentiation and function |
| Osteoblasts | PCS and IS | Decreased cell viability and cell proliferation and increased ROS production |
| IS | Decreases PTH receptor expression | |
| Promotes apoptosis | ||
| PAA | Inhibits proliferation and differentiation |
PAA, phenylacetic acid; IAA, indole acetic acid; IS, indoxyl sulfate; OAT, organic acid transporters; PC, p-cresol; PCS, p-cresyl sulfate; PTH, parathyroid hormone; RAS, renin–angiotensin system; ROS, oxygen free radicals; TMAO, trimethylamine N-oxide.
In recent years there is a growing interest in restoring the symbiosis of intestinal microflora in CKD aiming to reduce the generation of uraemic toxins, oxidative stress, and inflammation.79
- a)
High fibre diet: a high fibre diet increases the production of SCFA, which provides energy to the intestinal flora and allows amino acids that reach the colon to be incorporated into bacterial proteins and be excreted instead of being fermented into uraemic solutes. In addition, SCFAs are used as substrate by the intestinal mucosa helping to maintain their functionality and integrity. Fibre increases intestinal transit reducing the time for fermentation of amino acids and improves the composition of microflora which reduces the production of undesirable solutes. In CKD patients, there is a direct relationship between dietary protein/fibre ratio and PCS and IS levels, so a diet with a low protein/fibre ratio should be beneficial.80 In healthy subjects, a vegetarian diet, as compared with the omnivore diet, reduces the generation IS or PCS; this effect was related to the higher fibre and lower protein content of the vegetarian diet.81 A very low protein diet (0.3g/kg body weight/day) supplemented with amino acid keto-analogues also reduces IS levels in patients with CKD.82
Several therapeutic interventions have recently explored to improve the dysbiosis of the intestinal microflora, reduce the absorption of uraemic toxins and the passage of endotoxins from the intestinal lumen.
- b)
Prebiotics, probiotics, and symbiotics: the generation of uraemic toxins could be reduced by selectively increasing saccharolyticbacteria (which digest dietary fibre) and decreasing proteolytic bacteria (protein and amino acid fermenters) in the colon. The main regulator of metabolism of colon bacteria is the availability of nutrients and specifically the rate of fermentable carbohydrates vs. nitrogen.
Prebiotics are non-digestible food components which, through selective fermentation, allow for specific changes in the composition or activity in gastrointestinal microflora, which are beneficial to the health and well-being of the host. Prebiotics stimulate the growth or activity of one or a limited number of bacteria in the colon; they may increase carbohydrate fermentables vs. nitrogen; they include inulin, fructooligosaccharides, galactooligosaccharides, etc. Inulin enriched with oligofructose reduces the generation of PCS and the serum concentrations in hemodialysis patients, but has no effect on IS.83 Resistant starch reduces IS levels in hemodialysis patients and reduces PCS but not significantly.84 In a CKD rat model, a diet rich in resistant starch delayed the progression of CKD and attenuated oxidative stress and inflammation.85 Currently, a randomised, crossover, double-blind, phase 2 clinical trial in patients with stage 3b-4 CKD is examining the effect of the supplementation of arabinoxylan-oligosaccharides on plasma levels of PCS and indole derivatives urinary excretion of these compounds and insulin resistance are also examined.86
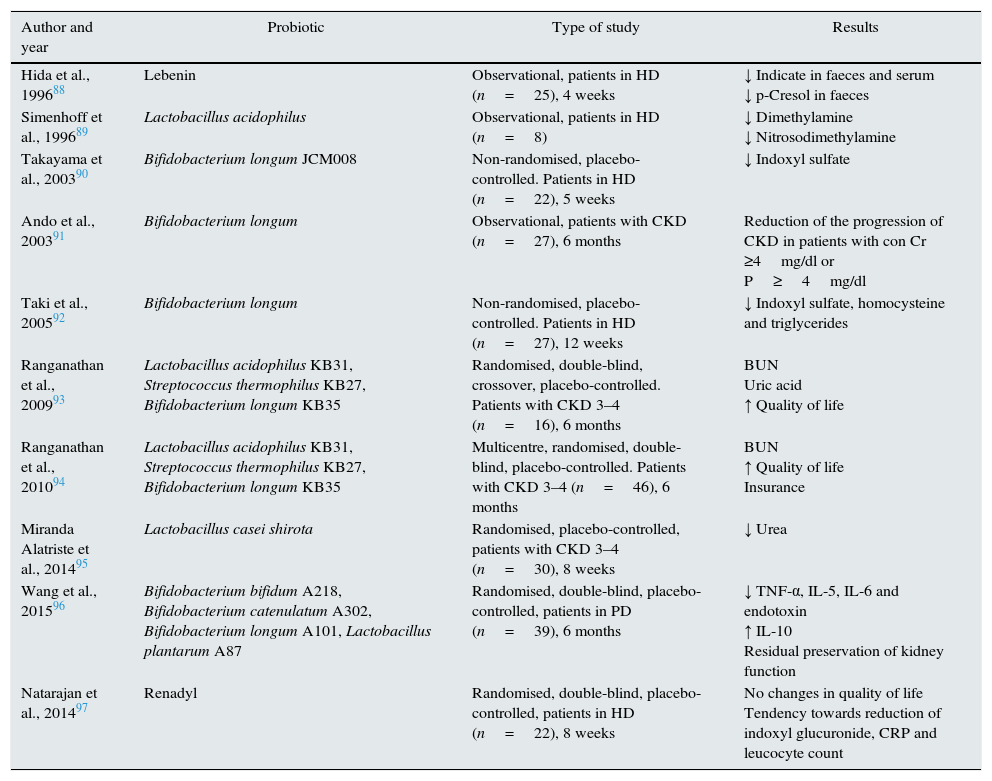
Probiotics are defined as “living micro-organisms” that, being administered in adequate amounts, provide a health benefit to the host. A recent review evaluates the potential benefits of probiotics in general and especially in CKD.87 The efficacy of probiotics to decrease levels of uraemic toxins and to delay the progression of CKD has been investigated in in vitro models, animal models and in patients with CKD. However, to date, there are no large-scale quality intervention studies and studies on clinical events to support their widespread use. There are only small studies, most of which,88,92–95 but not all,97 observe a decrease in uraemic toxin levels. Administration of Bifidobacterium longum in enteric capsules to patients with CKD had minimal effects on the progression of the disease in patients with CKD.91 However, a randomised, double-blind trial in patients on peritoneal dialysis observed a significant reduction in serum proinflammatory endotoxin and cytokine levels, an increase in serum IL-10 levels, and the preservation of residual renal function after 6 months of treatment with a probiotic96 (Table 4).
Table 4.Clinical studies with probiotics in patients with CKD and their effects.
Author and year Probiotic Type of study Results Hida et al., 199688 Lebenin Observational, patients in HD (n=25), 4 weeks ↓ Indicate in faeces and serum
↓ p-Cresol in faecesSimenhoff et al., 199689 Lactobacillus acidophilus Observational, patients in HD (n=8) ↓ Dimethylamine
↓ NitrosodimethylamineTakayama et al., 200390 Bifidobacterium longum JCM008 Non-randomised, placebo-controlled. Patients in HD (n=22), 5 weeks ↓ Indoxyl sulfate Ando et al., 200391 Bifidobacterium longum Observational, patients with CKD (n=27), 6 months Reduction of the progression of CKD in patients with con Cr ≥4mg/dl or P≥4mg/dl Taki et al., 200592 Bifidobacterium longum Non-randomised, placebo-controlled. Patients in HD (n=27), 12 weeks ↓ Indoxyl sulfate, homocysteine and triglycerides Ranganathan et al., 200993 Lactobacillus acidophilus KB31, Streptococcus thermophilus KB27, Bifidobacterium longum KB35 Randomised, double-blind, crossover, placebo-controlled. Patients with CKD 3–4 (n=16), 6 months BUN
Uric acid
↑ Quality of lifeRanganathan et al., 201094 Lactobacillus acidophilus KB31, Streptococcus thermophilus KB27, Bifidobacterium longum KB35 Multicentre, randomised, double-blind, placebo-controlled. Patients with CKD 3–4 (n=46), 6 months BUN
↑ Quality of life
InsuranceMiranda Alatriste et al., 201495 Lactobacillus casei shirota Randomised, placebo-controlled, patients with CKD 3–4 (n=30), 8 weeks ↓ Urea Wang et al., 201596 Bifidobacterium bifidum A218, Bifidobacterium catenulatum A302, Bifidobacterium longum A101, Lactobacillus plantarum A87 Randomised, double-blind, placebo-controlled, patients in PD (n=39), 6 months ↓ TNF-α, IL-5, IL-6 and endotoxin
↑ IL-10
Residual preservation of kidney functionNatarajan et al., 201497 Renadyl Randomised, double-blind, placebo-controlled, patients in HD (n=22), 8 weeks No changes in quality of life
Tendency towards reduction of indoxyl glucuronide, CRP and leucocyte countCr, creatinine; PD, peritoneal dialysis; CKD, chronic kidney disease; HD, haemodialysis; IL, interleukin; P, phosphorus; CRP, C-reactive protein.
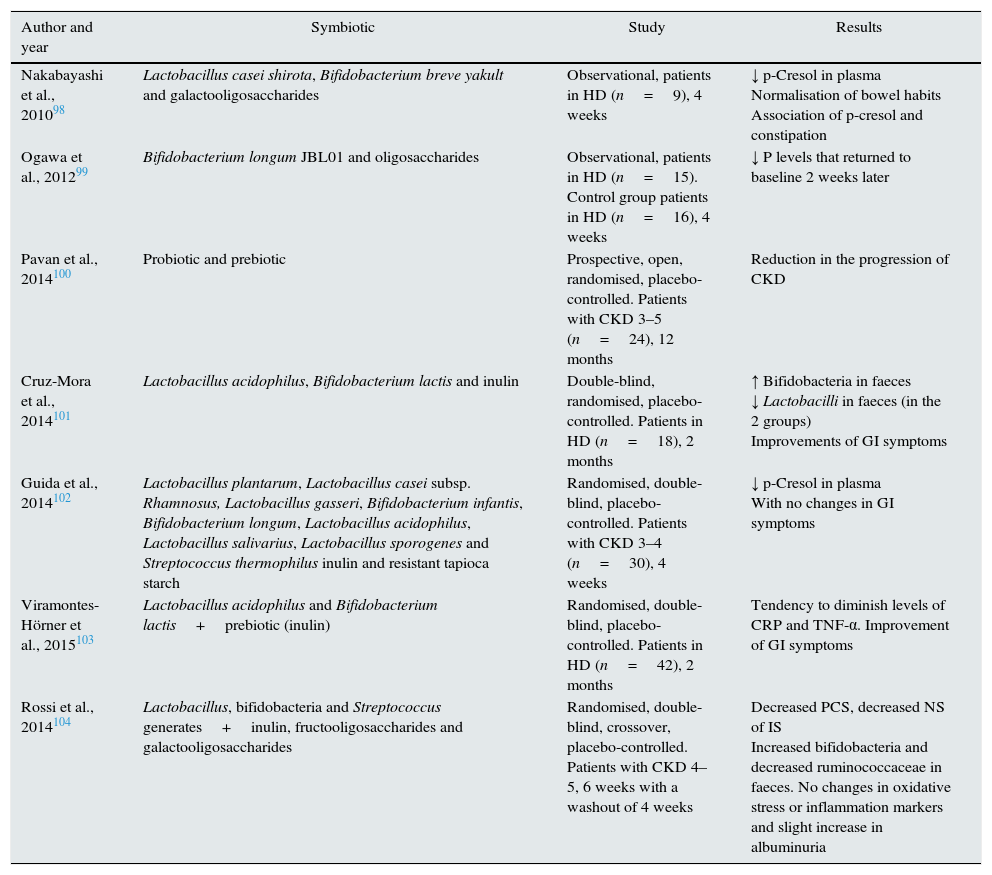
Symbiotics are probiotic supplements combined with prebiotics. In hemodialysis patients, on treatment with a symbiotic there is a decrease in the level of PCS, but not those of IS,98 which was confirmed in another study.102 Another study observed a delay in the progression of CKD with symbiotic treatment,100 while another study did not observe a significant improvement in inflammation markers.103 Finally, a randomised, double-blind, crossover study in patients with CKD104 demonstrated a reduction in PCS levels, a non-significant decrease in IS and an increase in bifidobacteria and reduction of faecal ruminococcus, but no change in inflammation markers, oxidative stress, or endotoxins, although a slight increase in albuminuria was observed (Table 5).
Table 5.Clinical studies with symbiotics in patients with CKD and their effects.
Author and year Symbiotic Study Results Nakabayashi et al., 201098 Lactobacillus casei shirota, Bifidobacterium breve yakult and galactooligosaccharides Observational, patients in HD (n=9), 4 weeks ↓ p-Cresol in plasma
Normalisation of bowel habits
Association of p-cresol and constipationOgawa et al., 201299 Bifidobacterium longum JBL01 and oligosaccharides Observational, patients in HD (n=15). Control group patients in HD (n=16), 4 weeks ↓ P levels that returned to baseline 2 weeks later Pavan et al., 2014100 Probiotic and prebiotic Prospective, open, randomised, placebo-controlled. Patients with CKD 3–5 (n=24), 12 months Reduction in the progression of CKD Cruz-Mora et al., 2014101 Lactobacillus acidophilus, Bifidobacterium lactis and inulin Double-blind, randomised, placebo-controlled. Patients in HD (n=18), 2 months ↑ Bifidobacteria in faeces
↓ Lactobacilli in faeces (in the 2 groups)
Improvements of GI symptomsGuida et al., 2014102 Lactobacillus plantarum, Lactobacillus casei subsp. Rhamnosus, Lactobacillus gasseri, Bifidobacterium infantis, Bifidobacterium longum, Lactobacillus acidophilus, Lactobacillus salivarius, Lactobacillus sporogenes and Streptococcus thermophilus inulin and resistant tapioca starch Randomised, double-blind, placebo-controlled. Patients with CKD 3–4 (n=30), 4 weeks ↓ p-Cresol in plasma
With no changes in GI symptomsViramontes-Hörner et al., 2015103 Lactobacillus acidophilus and Bifidobacterium lactis+prebiotic (inulin) Randomised, double-blind, placebo-controlled. Patients in HD (n=42), 2 months Tendency to diminish levels of CRP and TNF-α. Improvement of GI symptoms Rossi et al., 2014104 Lactobacillus, bifidobacteria and Streptococcus generates+inulin, fructooligosaccharides and galactooligosaccharides Randomised, double-blind, crossover, placebo-controlled. Patients with CKD 4–5, 6 weeks with a washout of 4 weeks Decreased PCS, decreased NS of IS
Increased bifidobacteria and decreased ruminococcaceae in faeces. No changes in oxidative stress or inflammation markers and slight increase in albuminuriaCRD, chronic kidney disease; GI, gastrointestinal; HD, haemodialysis; NO, nonsignificant; P, phosphorus; CRP, C-reactive protein; PCS, p-cresyl sulfate.
One of the major limitations of probiotic or symbiotic therapy is that no study has yet demonstrated the sustained survival of probiotics in the dysbiotic colon of patients with CKD. There are also no studies that have evaluated the effect of these treatments on the levels of TMAO in this population. In choosing probiotics, the contribution of urease-containing bacteria must be considered, since they may increase intestinal ammonia generation, which may damage epithelial tight junctions, and increase intestinal permeability to the passage of endotoxins from the intestinal lumen.33,34
- c)
Adsorptive therapies: the use of oral sorbents could decrease uraemic toxins and circulating intestinal endotoxins. AST-120 oral sorbents decrease IS levels in a dose-dependent manner.105 In addition, a reduction in the IS, PCS, or phenyl sulfate and oxidative stress levels have been described in patients on haemodialysis.106 Other authors have reported that the administration of AST-120 improves the erythropoietic response to CERA.107 AST-120 improves intestinal barrier dysfunction and decreases endotoxin plasma levels, inflammation markers and oxidative stress in a CKD model in rats.108
Although small randomised, controlled studies in experimental animals and retrospective studies in patients have indicated a nephroprotective effect of AST-120 (reviewed by Schulman et al.109), a subsequent large randomised, controlled trial in patients with CKD was unable to confirm this.109 The study had some methodological limitations, but also suggested the possibility that the objective of treating specific uraemic toxins may not be sufficient. However, another retrospective study of the long-term effects of AST-120 on patients with stage 3–5 CKD showed a reduction in the risk of progression to dialysis, mortality, cardiac events and vascular accident vs. those patients who did not receive it.110
Although a beneficial effect of sevelamer on IS and PCS has been described in in vitro studies, in vivo studies in mice or patients have not demonstrated a reduction in the levels of these uraemic toxins.111 However, sevelamer does reduce endotoxin levels and systemic inflammation in patients on haemodialysis.112,113
Key concepts- 1.
In CKD, there is a dysbiosis of the intestinal microflora.
- 2.
Intestinal microflora generate uraemic toxins that are absorbed and accumulate in CKD, and are associated with increased oxidative stress and inflammation.
- 3.
In CKD, there is an increase in the permeability of the intestinal barrier that allows the passage into the systemic circulation of endotoxins and other bacterial products that aggravate the inflammatory state of CKD.
- 4.
Changes in diet composition could improve microflora dysbiosis in CKD, reduce uraemic toxin levels, or restore intestinal mucosal permeability in CKD patients.
- 5.
The use of probiotics, prebiotics or symbiotics opens an alternative in the treatment of intestinal dysbiosis associated with CKD, and may play a role in slowing the progression of CKD and in preventing relevant associated complications such as mortality and cardiovascular risk.
The authors have no conflicts of interest to declare.
This work was supported by grants from Fondo de Investigaciones Sanitarias (PI16/01298), Madrid Society of Nephrology and REDinREN.
Please cite this article as: Guldris SC, Parra EG, Amenós AC. Microbiota intestinal en la enfermedad renal crónica. Nefrología. 2017;37:9–19.



